Chapter 10. Habitat Fragmentation and Connectivity
Authors:
Karl Didier, Wildlife Conservation Society
Janice Thomson, The Wilderness Society
Contributors:
Silvio Frosini De Barros Ferraz, University of São Paulo, Brazil
Ben White, Global Land Cover Facility
Reviewers:
Kurt Riitters, United States Forest Service
Peter Vogt, Joint Research Centre
Valerie Kapos, UNEP World Conservation Monitoring Centre
Remote sensing based indicators for fragmentation and connectivity:
- Total number of land cover types
- Patch size (largest, average)
- Patch density
- Perimeter-to-area ratio (average)
- Core area index
- Fractal dimension
- Distance to nearest neighbor (average)
- Contagion
- Juxtaposition index
- Road length
- Road density
10.1 Background
Habitat fragmentation is the division of ecosystems or habitats into smaller, less connected patches. It has been defined as the “creation of a complex mosaic of spatial and successional habitats from formerly contiguous habitat” (Lehmkuhl and Ruggiero 1991). Fragmentation results from both natural and human-caused processes.
Connectivity and intactness are closely related concepts (D’Eon et al 2002) to fragmentation: an area that is not fragmented provides continuous habitat, and an area that is fragmented is poorly connected. One can envision a continuum of intactness from a pristine environment on one end to a totally developed environment on the other, with fragmented landscapes occupying the middle of the continuum.
Habitat fragmentation is the result of a number of natural processes, including fire and windthrow (CBD 2005). However, in the context of biodiversity monitoring, fragmentation usually refers to anthropogenic causes such as logging, agricultural development, urban development, and infrastructure development (e.g., roads, utility corridors, oil and gas development, and irrigation canals). Ideally, a baseline condition is established so that an understanding of naturally occurring habitat fragmentation and connectivity from natural processes in a given area or ecosystem type is achieved. Then fragmentation and connectivity indicators are measured to determine human-induced changes that have occurred within the landscape.
There is wide scientific consensus that fragmentation, when coupled with habitat loss, has had overall negative impacts on biodiversity, in terms of both the total number of species in an area and abundance within individual species (e.g., Lovejoy et al. 1986; Laurance et al. 1997; and Nepstad et al. 1999). Many scientists consider the destruction and fragmentation of natural habitats to be a leading cause of the decline and loss of native species (Harris 1984; Wilcox and Murphy 1985; Pickett and White 1985; Wiens et al. 1985; Wilcove 1989; Turner et al. 1993; Noss and Cooperrider 1994; Reice 1994; Newmark 1995; Sinclair et al. 1995; Soule and Terborgh 1999). Negative impacts of fragmentation include decreases in mobility and dispersal of organisms and other natural resources (water, nutrients), increases in mortality resulting from increased exposure to threats (e.g., hunting, fire), and increases in competition from nonnative species.
Roads are a widespread threat to biodiversity loss around the globe through habitat fragmentation. However, roads are also a needed human infrastructure for subsistence activities, trade and economic development, education, medical services, recreation, and other activities of societies. The need for human access should be balanced against the ecological impacts, including reduction in core habitat areas for wildlife; loss of connectivity for wildlife; increase in patch edge effects; diminished animal use of habitats because of noise, dust emissions, and increased presence of humans; interference with wildlife life-history functions (e.g., courtship, nesting, and migration); changes in human behavior such as increased poaching, unethical hunting practices, and recreational activities; physical changes such as the degradation of aquatic habitats through alteration of stream banks and increased sediment loads, spread of exotic species, and mortality from collision with vehicles (Franklin and Forman 1987; Lehmkuhl and Ruggiero 1991; Reed et al. 1996; Forman and Alexander 1998; Trombulak and Frissell 2000). Construction of roads is often the catalyst for a dramatic increase in other anthropogenic threats to biodiversity, such as colonization, land-clearance, and invasion by exotic species.
Similarly, agricultural conversion plays a vital role for society while causing biodiversity loss through removal and fragmentation of natural ecosystem areas and replacement of species assemblages with exotic or domesticated taxa, and through degradation of remaining areas by factors associated with the new land use. For example, modern agricultural techniques can affect biodiversity off-site by (a) diverting water supplies (decreasing the area and/or quality of aquatic and open water habitats); (b) producing high-nutrient agricultural effluent; (c) causing soil erosion; (d) introducing and spreading invasive species, and (e) expanding edge habitats dominated by less threatened species. In addition, agricultural practices may introduce new processes into the disturbance regimes of natural areas (e.g., fire in tropical moist forest ecosystems) that further fragment landscapes.
The effects of fragmentation from roads, agriculture and other development extend well beyond their physical footprint (Franklin and Forman 1987; Chen et al. 1995; Lehmkuhl and Ruggiero 1991; Reed et al. 1996; Lyon and Christensen 2002; Lutz et al. 2003). For example, the openings caused by roads in forested landscapes has been shown to change microclimate conditions (i.e., increased evaporation, temperature, and solar radiation and decreased soil moisture) for some distance into the forest (Chen et al. 1995). Increased competition may occur for species that prefer edges or openings created by roads (Lehmkuhl and Ruggiero 1991). In landscapes fragmented by agriculture, natural habitat edges have been shown to degrade through time because of increased temperature, decreased humidity, increased wind shear, and other factors. As this edge relaxation continues, spaces that were previously occupied by native flora are subsequently colonized by species from adjacent agricultural or secondary ecosystems (Laurance et al. 1997).
Measures of fragmentation and connectivity are most useful as indicators when they can be linked to specific negative impacts on a particular species, habitat type, aquatic system, or other resource (e.g., measures of road density have been linked to reduced habitat effectiveness for elk by Lyon 1983). The response of individual organisms or biodiversity as a whole (e.g., species richness) to most fragmentation indicators requires further research to ensure that the indicators are useful for the system or species of interest. Only a few field studies have documented the negative relationship between connectivity indicators and species diversity (e.g., Goparaju et al. 2005; Jha et al. 2005). Review papers and literature reviews have been published compiling the scientific studies of the effects of roads (including fragmentation effects) on ecosystems and wildlife (Forman and Alexander 1998; Trombulak and Frissell 2000; Gucinski et al. 2001; Gains et al. 2003; Wyoming Game and Fish Department 2004). As a point of clarity, landscape fragmentation or intactness measures, while valuable as indicators, cannot measure all facets of ecosystem integrity. For instance, the “empty forest syndrome,” in which the canopy structure remains but the wildlife has been hunted out, cannot be effectively monitored through remote sensing and fragmentation metrics.
10.2 Potential role for remote sensing
Remote sensing may be used to map habitat types or land cover across a landscape and to map the impacts of natural and human-caused processes causing fragmentation (e.g., cultivated land or fire scars). The very same imagery and classifications used to determine trends in habitats and ecosystems can be reused for fragmentation or connectivity analyses. In addition to land cover classes, categories of habitat vs. nonhabitat may also be used. Mapped results become the base data for calculating landscape fragmentation metrics and are relatively easy to generate using existing software packages (e.g.,
FRAGSTATS or
Patch Analyst).
Alternatively, the causes of fragmentation (e.g., roads, agriculture, or fire) can be mapped, and then the reciprocal of these areas can be classified as intact patches of habitat. For example, the location of agricultural land use is traceable through time by a wide variety of space-based remote sensing imagery and is currently being quantified annually at local, regional, and national scales for selected parts of the globe (Loveland et al. 1995; Liang et al. 2004; Kerr and Cihlar 2003; Senay and Elliot 2002). At a continental scale, advanced very high resolution radiometer (
AVHRR) and
MODIS satellite systems can be used to assess both the spatial extent and the health of agricultural systems. Specific types of agricultural systems and different crops can be identified using higher resolution TM, ETM+, and IRS satellites (Oetter et al. 2002). Data from these moderate resolution sensors can also be used in local or site-based analyses for evaluating how an agricultural site is integrated spatially within an agricultural/natural habitat mosaic.
The spatial pattern of roads can be generated from moderate- to high-resolution image data and then used as an overlay to create habitat patches. Weller et al. (2002) generated road data from digital air photos to calculate fragmentation metrics as indicators of wildlife impacts. Moderate-resolution sensors such as TM, SPOT, and IRS are use to delineate road systems and cover larger areas more quickly and cheaply. However, the drawback is that their coarser resolution will cause the omission of some roads that exist on the ground, depending upon road width, canopy type, and spectral contrast. Many of these problems are overcome by using high-resolution photos and digital sensors, typically 1–4 metres in resolution (such as air photos,
IKONOS, and
QuickBird). Images from these sensors allow direct spatial recognition of the roads in many cases and require less spectral contrast between the road and the surrounding landscape. Drawbacks to these sensors include the high image cost per unit area and the substantially larger volume of data required to cover a project area. In most cases, regional or national projects with high-resolution data sets are not practical at this time because of cost and time required for analysis.
The delineation of roads has typically been done by operator visual interpretation and on-screen digitizing. A human operator can reasonably bring the necessary knowledge and recognition of shape and context in an image; however, techniques are being developed to automate the process of road delineation. It is still associated with a high degree of error and typically still requires a human operator to refine the final road product (Vosselman 1996).
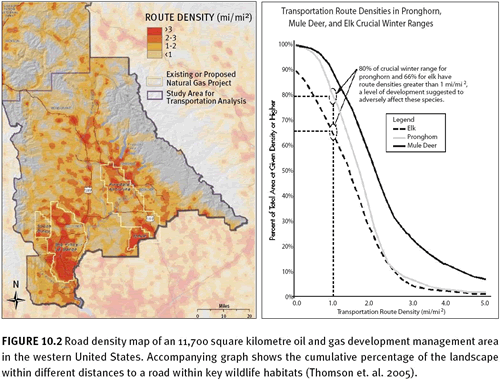
Weller et al. (2002) describes the use of digital air photos (1-metre pixels) to digitize roads in the rangeland in the western United States. (See figure 10.1) The authors digitized roads in an oil and gas field and calculated metrics for the total length of road, road density (the average for the study area and density variation across the landscape), road effect zones, and core habitat areas. Fragmentation metric results were compared with biological literature linking specific road metrics or indicators to the impacts on specific species. For example, the average road density was more than 8 times higher than the road density suggested to adversely affect pronghorn (
Antelocarpa americana), a prominent big game species that winters in the area.
A similar report by Thomson et al. (2005) measures a number of indicators, including road density, core area, and cumulative distance to road values. The measures are made for the full landscape and for specific land management units, for oil and gas development units, and for wildlife habitat units to allow for the evaluation of current conditions for wildlife and to serve as a baseline for future management options. Comparing indicator results to the wildlife literature, the authors found that 66 percent of elk winter range has road densities greater than what may eliminate effective habitat (figure 10.2). In addition, 100 percent of sage grouse breeding and rearing areas are closer to a road than the avoidance distance recommended during breeding season.
10.3 Connectivity in the marine environment
Remotely sensed imagery can also be used to measure some types of connectivity in marine environments. Many species depend on specific habitats (e.g., mangroves, seagrasses, and coral reefs) or networks of these habitats for one or more of their life stages (e.g., Phinney et al. 2001). In addition, the physical condition of channels or inlets can be monitored with imagery. These habitats and the changes in these habitats (depending on size) are often visible using Landsat, ASTER, and MODIS. Many species thrive in productive areas; thus, imagery that provides information about ocean productivity (Ocean Color from
SeaWiFS) can let one separate patches of habitat from nonhabitat. Tracking this through time (years), one could potentially see whether areas that are historically productive are still productive or whether these patterns are changing because of pollution or global warming. Connectivity can also be measured in the ocean using altimetry data (e.g.,
TOPEX) to examine current speed and direction, which can illustrate whether sites are upstream or downstream in terms of larval (and threats) dispersal.
10.4 Ecological and biodiversity relevance
Some studies have directly measured the relationship between fragmentation metrics and the response of biodiversity, but generally only for single species or small groups at relatively fine spatial scales (e.g., Chust et al. 2004; Homan et al. 2004; and Jacuemyn et al. 2002). Few studies have directly linked fragmentation metrics to the response of biodiversity as a whole (abundance or richness), although modelling attempts have been made to generalize the impacts of fragmentation (e.g., Tischendorf 2001; D’Eon et al. 2002).

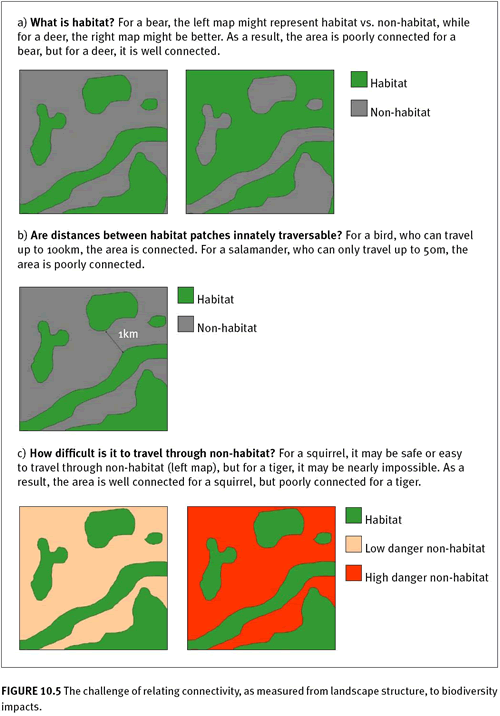
The causes of fragmentation, as well as biological connectivity requirements (D’Eon et al. 2002) should form the basis for selection of the appropriate fragmentation/connectivity indicators. Whether a landscape is fragmented or connected depends highly on which particular natural functions of the landscape are in peril. An area can be well or poorly connected from the perspective of water flow, soil organisms, nutrients, terrestrial wildlife, dispersing seeds, people, and a myriad of other elements. Any change in fragmentation could have wildly different impacts on these various elements of that landscape. For example, figure 10.5 considers patch connectivity of a landscape (i.e., a configuration metric) from the viewpoint of different organisms. Depending on the particular species, the landscape could be well connected or completed fragmented (Tischendorf and Fahrig 2000; Tischendorf 2001). In addition, if one were considering fragmentation of the landscape for nonliving elements, such as the movement of water, the context would be completely different, and the selection of an indicator that measures habitat connectivity (e.g., nearest neighbor distance) may be irrelevant.
Most fragmentation studies combine the effects of habitat loss on biodiversity with the effects of fragmentation. It is well established that habitat loss has consistently negative effects on biodiversity. Once effects of fragmentation are decoupled from habitat loss, however, fragmentation has been shown to have a weaker effect on biodiversity (although recently Koper et al. (2007) have called this consensus view into question). In many cases it may even have a positive effect (see Fahrig 2003 for more discussion and multiple references.) Therefore, in most cases habitat or ecosystem loss should be considered a superior predictor — and indicator — of biodiversity loss. However, in specific cases where habitat needs of focal species are well understood, fragmentation and/or connectivity should be considered in addition to habitat extent. In other cases, it is useful simply to clarify the relative concentration or diffusion of habitat loss (or lack thereof). Given the ease with which fragmentation information can be produced when input data such as land cover already exists, it can easily be produced as a trial indicator.
10.5 Assessment at national or continental scales
It is theoretically possible to develop a national indicator of connectivity that represents the range of biodiversity. Furthermore, index or indicator calculations can be accomplished cheaply and quickly if using data from a pre-existing land cover change series. However, to truly be useful in a management sense, the indicator would have to incorporate measurements across a range of spatial scales, and be interpreted separately for a number of different species with different habitat requirements.
An important consideration is that the accuracy of remotely sensed fragmentation metrics have not been verified using ground truth information, partly because the metrics are derived from land cover data sets. These metrics are susceptible to error since the land-cover data sets on which they are usually based already contain some level of error. Overall accuracy of land cover classifications are are often 80% or lower. However, fragmentation error might exceed that of land cover, because fragmentation is concerned with the edge characteristics of patches and edges, which are where most of the errors in land cover classifications occur (O’Neill et al. 1999). Furthermore, measuring trends requires comparable datasets spanning the desired time period. In most instances, comparable land cover classifications at the scale of desired study are not available. Therefore, most fragmentation studies are snapshots, rather than depictions of trends.
Figure 10.3 represents a national study of change in forest loss and fragmentation over a period of fifty years.
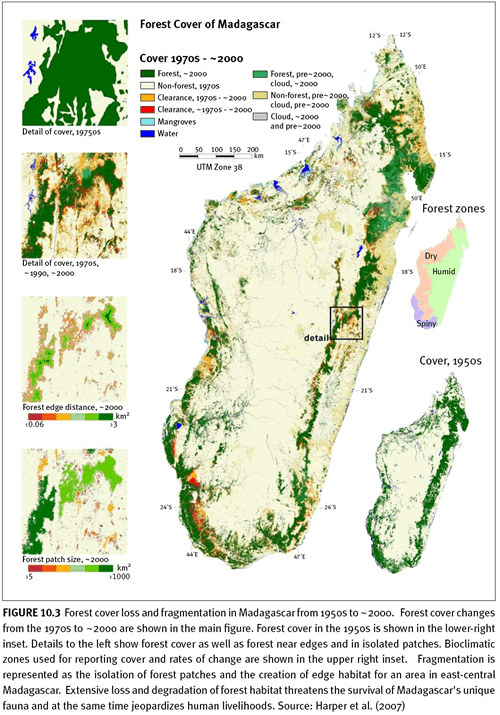
Madagascar's forests are among the most biologically rich and unique in the world. Past estimates of forest cover and deforestation have varied widely, so Harper et al. (2007) measured deforestation and forest fragmentation in Madagascar from the 1950s to ~2000 using aerial photography and Landsat imagery. Forest cover decreased by almost 40 percent from the 1950s to ~2000, with a reduction in 'core forest' more than one kilometer from a non-forest edge of almost 80 percent. This forest destruction and degradation threatens thousands of species with extinction. Country-wide coverage of high-resolution, validated forest cover and deforestation data enables the precise monitoring of trends in habitat extent and fragmentation critical for assessment of species' conservation status.
On a continental scale, the work of Riitters et al. (2000) illustrates a study measuring forest fragmentation across continents for a global view of changing forest patterns using AVHRR 1-kilometre land cover data. They developed a method for estimating perforated, edge, and transitional habitats, important indicators in progressively fragmenting forest areas. (Figure 10.4 shows the results for Eurasia.)

More recent and improved methods for calculating perforated edge have been produced by Vogt et al. (2007a, b). Riitters et al. (2000) and Wade et al. (2003) describe global forest fragmentation as well. Such analyses have not yet been produced for other biomes (e.g., grasslands, savannas or deserts).
10.6 Fragmentation/connectivity metrics
At this time, there is no scientific consensus as to which individual or set of indicators best represents the impacts of fragmentation on biodiversity as a whole (Calabrese and Fagan 2004). Hundreds of potential indicators of connectivity, based on the observed structure of the landscape, have been developed, and a number of software packages help users produce and explore the variety of indicators, including
FRAGSTATS (McGarigal and Marks 1995, McGarigal et al. 2002),
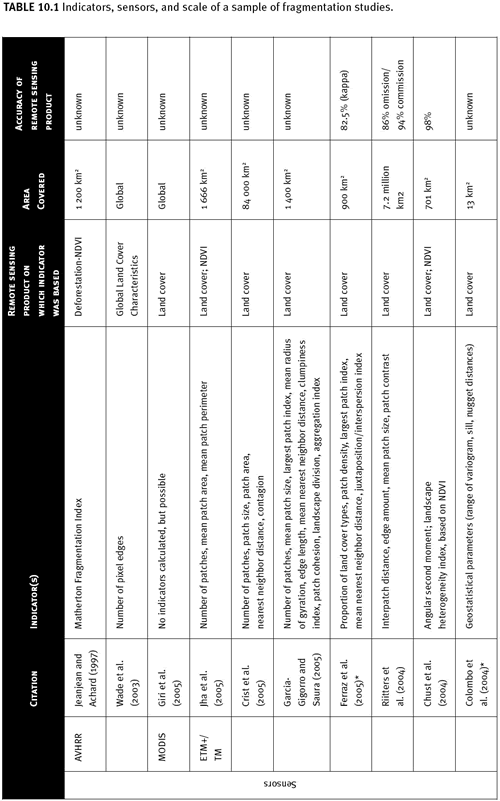
Patch Analyst Extension for ArcView, r.le for GRASS, and Leap II. However, ease of calculation, relationship to the biology of organisms (e.g., dispersal capability), and consistency across spatial scales of fragmentation indicators vary greatly (Tischendorf 2001; Rutledge 2002). Fragmentation/connectivity metrics can be measured for individual patches (e.g., patch size) or aggregated across patches at broader scales (e.g., mean patch size within an area). Examples of research using a range of image data and many different types of metrics as indicators are given in Table 10.1. Here, we restrict ourselves to a discussion of metrics meant to characterize broad spatial scales, often referred to as “landscape metrics” (Fahrig 2003). We also bias our discussion to metrics that can be easily measured and compared over two to three time periods in an assessment of trends, as opposed to metrics to compare between large regions or nations.

There are several review papers of fragmentation metrics that categorize and review the utility of landscape metrics (e.g., Garicia-Gigorro and Saura 2005; Calabrese and Fagan 2004; Moilanen and Nieminen 2002; and Rutledge 2002). Here, we follow a relatively simple and easily interpretable framework for categorizing metrics into five groups, corresponding to different ways that fragmentation is manifest within a landscape: (1) composition, (2) patch size/density, (3) shape, (4) configuration of landscapes, and (5) route networks. However, existing metrics do not always fit neatly into these categories, because often they correspond to two or more characteristics of fragmentation. Composition metrics are those that describe the types of patch (e.g., forest, agriculture, urban, or habitat versus nonhabitat), including patch-type richness, evenness, and diversity indices (e.g., Shannon’s Index). A simple, but consistently useful metric (Cain et al. 1997) suggested by O’Neill et al. (1999) is simply the total number of land cover types (richness). Places or landscapes that are becoming more fragmented over time would see an increase in the number of land cover types.
Patch size/density metrics are sometimes listed as part of composition metrics, but are meant to characterize the size or number of habitat patches within an area. Patch size metrics come in varying forms, from the simple patch density to the more complex effective mesh size (Jaeger 2000). Riitters et al. (2004) used mean patch size. Places or landscapes exhibiting more fragmentation over time would show a decrease in mean patch size.
Shape metrics quantify the complexity of patches, from patches that are more compact (circular) to patches that are more complex in shape. Measures of shape generally are for some perimeter-to-area ratios, but also include more species-specific measurements such as core area index (McGarigal 2002). Fractal dimension is also another common measurement of shape. O’Neill et al. (1999) suggested using two types of shape measurement—average perimeter-area ratio and fractal dimension—for regional assessments, corresponding respectively to different interpretations of patch shape and patch compaction, as described by Riitters et al. (1995). Places or landscapes becoming more fragmented over time would see an increase in the mean perimeter-area ratio and fractal dimension.
Configuration metrics more formally characterize the degree of connectivity or isolation among habitat patches. Metrics based on the distance to nearest neighbor(s) are very common, but range from simple to difficult to calculate based on which patches are considered neighbors. Contagion (raster-based) is a commonly used metric that measures adjacency (the degree of adjacency of pixels across a landscape), and juxtaposition index (McGarigal and Marks 1995) is its vector-based cousin. Unlike contagion, Frohn’s (1997) patch-per-unit-area index allows consistent comparison across different analysis extents and resolutions. Riitters et al. (2004) used interpatch distance (average nearest neighbor distance) to measure fragmentation in U.S. forests. O’Neill et al. (1999) suggests calculating a version of contagion for regional-scale assessments. Route network metrics are used to characterize the fragmentation caused by roads, utility corridors, or other linear features that break up otherwise contiguous habitat patches. Road length and road density are among the most common metrics. The area or percent area of habitat within threshold distances of roads (or cumulative distribution for all distances) is also used for specific ecological impacts occurring within known distances of roads. By using the route network data to segment the habitat data, one can use many of the patch metrics described above.
10.7 Key considerations for selecting metrics
No single fragmentation metric (Betts 2000) or even a small set of metrics is perfect for every application. No single metric can represent real-world fragmentation in all its varied forms. To represent the different possible ways that fragmentation manifests itself, a group of at least four to five metrics should be selected to characterize patch composition, size, shape, and configuration across broad areas. As mentioned earlier, specific connectivity needs of local biodiversity should help narrow down the choice of metrics. Further selection of the specific metric should be made with reference to the following considerations.
 10.7.1 Sensitivity to scale
10.7.1 Sensitivity to scale Many of the indicators listed above are sensitive to changes in the extent over which they are calculated and, particularly, to the resolution of the input data (e.g., Lawler et al. 2004). If the analysis extent and resolution are kept consistent, different areas or different points in time can be compared relatively easily, using many indicators. Further, O’Neill et al. (1999) suggests that over the range of scales encountered in typical remote sensing data (e.g., 10–100-metre resolution), most of indicators are relatively insensitive to resolution (see Wickham and Riitters 1995 for further discussion).
10.7.2 Calculation complexity and ease of interpretation The available indicators (e.g., in
FRAGSTATS) vary greatly in their calculation complexity and therefore also in the processing time needed to calculate them (and often their ease of interpretation). For instance, for comparing patch size between areas or points in time, mean patch size is likely to be easier to calculate and interpret than mesh size. Consideration of complexity as it relates to processing time will be affected by both the analysis extent and the resolution of the input data. Indicators based on pixel data are likely to require less processing and calculation time than those based on vector data. Indicators are most useful when they can be connected to specific resource needs (e.g., patch size requirements for certain species or road density thresholds for watershed aquatic integrity).
Case Study 10.1: Measuring Tropical Forest Fragmentation in the Amazon (Summary from Ferraz et al. 2005)
Indicators: the proportion of each land-use class, patch density, largest patch index, the mean nearest neighbor distance, and the interspersion/juxtaposition index
Potential monitoring scales: regional and national
Sensor: Landsat TM and ETM+
Imagery cost : Free from various institutions
a. Introduction The loss of tropical forests in the Amazon has been well documented, yet the landscape patterns and dynamics are not well studied or understood. Central Rondonia in Brazil is among the most heavily deforested areas within the Amazon. Forest is lost from small temporary clearings by small farmers and loggers and larger-scale forest removal for crops and pasture. These activities lead to a complex pattern of forest degradation, including nonforest patches and forest patches at different growth stages. This study uses satellite data from every other year from 1984 to 2002 to systematically measure changes in forest fragmentation over time to answer some of the following questions: “How did fragmentation occur? What are the landscape trends? When did the matrix transition from forest to pasture? What is the proportion of mature forest that represents a critical threshold? Would the landscape structure be sustainable with the implementation of the ‘permanent preservation area’ law (Brazil, 1965)?” The analysis of forest fragmentation described here is an integral part of a larger study of landscape dynamics in central Rondonia, including land cover mapping, land-use transitional probabilities, and evaluation of the sustainability of land-use changes in future scenarios.
b. Methods The study area covers a watershed of approximately 900 square kilometres. Landsat TM and ETM+ images were acquired in even-numbered years from 1984 through 2002. Images were provided by the Land-Use Dynamics Project of the Brazilian Institute for Space Research and by the Tropical Rain Forest Information Center at Michigan State University. For each image date, the images were classified into the three most important cover types in the region: mature forest, secondary forest, and pasture. For each classification, five measures of fragmentation were calculated as indices of landscape structure: the proportion of landscape (proportion of each land-use class in the study area—a measure of landscape composition); patch density (the number of patches of each land-use class per unit area—a measure of dissection of patches); largest patch index (proportion of the landscape occupied by the largest patch of each land use—a measure of patch dominance); the mean nearest neighbor distance (the mean distance between patches of the same class—a measure of connectivity); and the interspersion/juxtaposition index (indicator of the adjacency of land-use categories—a measure of landscape configuration). Fragmentation measures were generated using FRAGSTATS 3.3 software (McGarigal et al. 2002).
c. Results The changes in fragmentation measures over time provide indicators of the forest structural dynamics resulting from logging and agriculture. Mature forest has been progressively converted to pasture. Mature forest was lost at an average rate of about 2 percent per year.
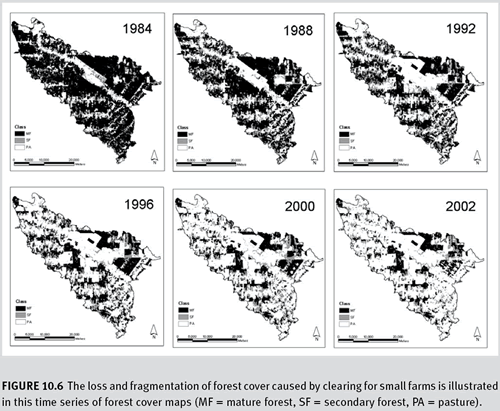
Figure 10.6 The loss and fragmentation of forest cover caused by clearing for small farms is illustrated in this time series of forest cover maps (MF = mature forest, SF = secondary forest, PA = pasture).
Looking at the frequency and size distribution of the forest patches showed specific trends in logging practices. Clear-cutting, converting mature forest to pasture, produced mostly small patches (about 1,000 0.5-hectare patches), but far fewer medium and large patches (100 1- to 50-hectare patches; 10 100- to 5,000-hectare patches). Selective logging, converting mature forest to secondary forest, occurs predominantly in small patches (5,000 patches smaller than 0.5 hectare). In fact, most of the total area logged occurs in patches smaller than 5 hectares.
Changing landscape structural dynamics are illustrated by trends in patch density, distance between patches, and the interspersion and juxtaposition index. The patch density for mature forest increased threefold from 1984 to 1996 and then remained relatively stable. The mean distance between patches of forest was little changed until 1996 and then approximately doubled by 2002. Distances between pasture patches followed the inverse pattern and dropped by about one quarter. Notably, 80 percent of deforestation occurs within 3 kilometres of a road, and 80 percent of clear-cutting occurs within 750 metres of existing pastures.
Authors also observed a critical fragmentation threshold of 35 percent mature forest cover. Mature forest loss occurred more rapidly as it declined below this value, suggesting that a minimum of 35 percent mature forest cover should be a target for natural resource managers.
10.8 Data and other resources
FRAGSTATS is a publicly available software package that computes a comprehensive variety of fragmentation metrics (McGarigal et al. 2002).
http://www.umass.edu/landeco/research/fragstats
FRAGSTATS-ARC commercial software integrates some of the most commonly used fragmentation metrics from FRAGSTATS into the widely used ARC/INFO GIS software. It provides a user-friendly interface familiar to many GIS professionals. This is a private product that may be purchased from the developer.
http://www.innovativegis.com/basis/present/fragstat
Patch Analyst is an extension for ArcView GIS software to analyse patches and their attributes. It can be obtained free of charge, but must be used under a license agreement because it is not in the public domain:
Patch Analyst
r.le programs is a public domain software that runs within GRASS GIS software. It contains a wide range of raster-based fragmentation metrics.
r.le programs
LEAP II uses an older version of FRAGSTATS that is designed to run under the Windows NT environment.
http://www.ai-geostats.org/
A broader description of all the software listed above can be found at:
fragstats links
Road-Mapping Tools: An issue of Photogrammetric Engineering and Remote Sensing (Vol. 70, No. 12, December 2004) has been devoted to the subject of linear-feature extraction from image data.
10.9 References
Amarnath, G., M.S.R. Murthy, S.J. Britto, G. Rajashekar and C.B.S. Dutt. 2003. Diagnostic analysis of conservation zones using remote sensing and GIS techniques in wet evergreen forests of the Western Ghats – An ecological hotspot, Tamil Nadu, India. Biodiversity and Conservation 12 (12): 2331-2359.
Betts, M. 2000. In search of ecological relevancy: a review of landscape fragmentation metrics and their application for the Fundy Model Forest. Working Group for Fundy Model Forest. New Brunswick Fish and Wildlife Research Units, University of New Brunswick, Canada.
http://www.unbf.ca/forestry/centers/fundy/metrics.htm
Cain, D.H., Riitters K., and K. Orvis. 1997. A multi-scale analysis of landscape metrics. Landscape Ecology 12:199-212.
Calabrese, J.M., and W.F. Fagan. 2004. A comparison-shopper’s guide to connectivity metrics Frontiers in Ecology and the Environment 2: 529-536.
CBD. 2005. Indicators for Assessing Progress towards the 2010 Target: Connectivity/Fragmentation of Ecosystems. Report from the tenth meeting of the Subsidiary Body on Scientific, Technical, and Technological Advice, Bangkok, Thailand.
Chen, J., J.F. Franklin, and T.A. Spies. 1995. Growing-season microclimatic gradients from clearcut edges into old-growth Douglas-fir forests. Ecological Applications 5(1): 74-86.
Chust, G., J.L. Pretus, D. Ducrot, and D. Ventura. 2004. Scale dependency of insect assemblages in response to landscape pattern. Landscape Ecology 19: 41-57.
Colombo, S., M. Chica-Olmo, F. Abarca, and H. Eva. 2004. Variographic analysis of tropical forest cover from multi-scale remotely sensed imagery. ISPRS Journal of Photogrammetry and Remote Sensing 58: 330-341.
Crist, M.R., B. Wilmer, and G.H. Aplet. 2005. Assessing the value of roadless areas in a conservation strategy: biodiversity and landscape connectivity in the northern Rockies. Journal of Applied Ecology 42: 181-191.
D'Eon, R., S.M. Glenn, I. Parfitt, and M. Fortin. 2002. Landscape connectivity as a function of scale and organism vagility in a real forested landscape. Conservation Ecology 6: 10.
Fahrig, L. 2003. Effects of habitat fragmentation on biodiversity. Annual Review of Ecology, Evolution, and Systematics. Vol. 34: 487-515
Ferraz, S.F.D., C.A. Vettorazzi, D.M. Theobald, and M.V.R. Ballester. 2005. Landscape dynamics of Amazonian deforestation between 1984 and 2002 in central Rondonia, Brazil: assessment and future scenarios. Forest ecology and management, 204: 67-83.
Forman, R.T., and L.E. Alexander. 1998. Roads and their major ecological effects. Annual Review of Ecological Systems 29: 207-231.
Franklin, J.F., and R.T.T. Forman. 1987. Creating landscape patterns by forest cutting: ecological consequences and principles. Landscape Ecology 1: 5-18.
Frohn, R.C. 1997. Remote sensing for landscape ecology: new metric indicators for monitoring, modeling, and assessment of ecosystems. Lewis Publishers, Boca Raton, Florida. 99p.
Fuller, D.O. 2001. Forest fragmentation in Loudoun County, Virginia, USA evaluated with multitemporal Landsat imagery. Landscape Ecology 16: 627-642.
Gaines, W., P. Singleton, and R. Ross. 2003. Assessing the Cumulative Effects of Linear Recreation Routes on Wildlife Habitats on the Okanogan and Wenatchee National Forests. General Technical Report PNW-GTR-586. U.S. Department of Agriculture, U.S. Forest Service, Pacific Northwest Research Station, Portland, OR.
Garcia-Gigorro, S., and S. Saura. 2005. Forest fragmentation estimated from remotely sensed data: is comparison across scales possible? Forest Science 51 (1): 51-63.
GeoCommunity. 2007. Vmap 0, Vector Map Level 0, description and data purchase. Available at:
http://store.geocomm.com/data/ (accessed May 2007)
Giri, C., Z.L. Ahu, and B. Reed. 2005. A comparative analysis of Global Land Cover 2000 and MODIS land cover data sets. Remote Sensing of Environment 94: 123-132.
Goparaju, L., A. Tripathi, C.S. Jha. 2005. Forest fragmentation impacts on phytodiversity – An analysis using remote sensing and GIS. Current Science 88: 1264-1274.
Gucinski, H., J.J. Furniss, R.R. Ziemer, and M.H. Brookes. 2001. Forest Roads : A Synthesis of Scientific Information. General Technical Report PNW-GTR-509. U.S. Department of Agriculture, U.S. Forest Service, Pacific Northwest Research Station, Portland, OR.
Harper, G.J., Steininger, M.K., Tucker, C.J., Juhn, D. and F. Hawkins. 2007. Fifty years of deforestation and forest fragmentation in Madagascar. Environmental Conservation, accepted, 2007.
Harris, L.D. 1984. The Fragmented Forest: Island Biogeography Theory and the Preservation of Biotic Diversity. University of Chicago Press, Chicago, IL.
Homan, R.N., B.S. Windmiller, and J.M. Reed. 2004. Critical thresholds associated with habitat loss for two vernal pool-breeding amphibians. Ecological Applications 14: 1547-1553.
Jacquemyn, H., R. Brys, & M. Hermy. 2002. Pathc occupancy, population size and reproductive success of a forest herb (Primula elatior) in a fragmented landscape.
Oecologia 130: 617-625.
Jaeger, J.A.G. 2000. Landscape division, splitting index, and effective mesh size: new measures of landscape fragmentation. Landscape Ecology 15: 115-130.
Jeanjean, H., and F. Achard. 1997. A new approach for tropical forest area monitoring using multiple spatial resolution satellite imagery. International Journal of Remote Sensing 18: 2455-2461.
Jha, C.S., L. Goparaju, A. Tripathi, B. Gharai, A.S. Raghubanshi, and J.S. Singh. 2005. Forest fragmentation and its impact on species diversity: an analysis using remote sensing and GIS. Biodiversity and Conservation 14: 1681-1698.
Kerr, J.T. and J. Cihlar 2003. Land use and cover with intensity of agriculture for Canada from satellite and census data. Global Ecology and Biogeography 12 (2): 161.
Koper, N., Schmiegelow, F.K.A., Merrill, E.H., 2007. Residuals cannot distinguish between ecological effects of habitat amount and fragmentation: implications for the debate. Landscape Ecology (online first) DOI 10.1007/s10980-007-9083-9.
Laurance, W.F., S.G. Laurance, L.V. Ferreira, J.M. Rankin-de Merona, C. Gascon, and T.E. Lovejoy. 1997. Biomass collapse in Amazonian forest fragments. Science 278: 1117-1118.
Lawler, J. J., O’Connor, R. J., Hunsaker, C. T., Jones, K. B., Loveland, T. R., & White, D. (2004). The effects of habitat resolution on models of avian diversity and distributions: a comparison of two land-cover classifications. Landscape Ecology, 19(5), 515-30.
Lehmkuhl, J. F., and L. F. Ruggiero, 1991. Forest fragmentation in the Pacific Northwest and its potential effects on wildlife. Pages 35-46 in: Ruggiero, L.F., K. B. Aubry, A. B. Carey, and M.H. Huff, tech. coords. Wildlife and Vegetation of Unmanaged Douglas-fir Forests. USDA Forest Service Gen. Tech. Rep. PNW-GTR-285. Portland, OR.
Liang, S., H. Fang, G. Hoogenboom, J. Teasdale and M. Cavigelli 2004. Estimation of crop yield at the regional scale from Modis observations. International Geoscience and Remote Sensing Symposium Proceedings 3: 1625- 1628.
Loveland, T.R., Merchant, J.W., Brown, J.F., Ohlen, D.O., Reed, B.C., Olson, P. and J. Hutchinson. 1995. Seasonal land-cover regions of the United States. Ann. Ass. Of American Geographers 85(2):339. doi:10.1111/j.1467-8306.1995.tb0189.x
Lovejoy, T.E., R.O. Bierregarrd, A.B. Rylands, J.R. Malcolm, C.E. Quintela, L.H. Harper, K.S. Brown, A.H. Powell, G.V.N. Powell, H.O.R. Schubart, and M.B. Hays. 1986. Edge and other effects of isolation on Amazon forest fragments. In M.E. Soule, editor. Conservation Biology: the science of scarcity and diversity. Sinauer Associates, Sunderland, Massachusetts, USA.
Luoto, M., Kuussaari, M., & Toivonen, T. (2002). Modelling butterfly distribution based on remote sensing data. Journal of Biogeography, 29, 1027-37.
Lutz, D.W., B.F. Wakeling, L.H. Carpenter, D. Stroud, M. Cox, D. McWhirter, S. Rosenstock, L.C. Bender, and A.F. Reeve. 2003. Impacts and changes to mule deer habitat. Pages 13-61 in: de Vos, J.C. Jr., M.R. Conover, and N.E. Headrick, Eds. Mule Deer Conservation: Issues and Management Strategies. Jack H. Berryman Institute Press, Utah State Universtiy, Logan, UT.
Lyon, L.J. 1983. Road Density Models Describing Habitat Effectiveness for Elk. Journal of Forestry 81 (9): 592-613.
Lyon, L.J. and A.G. Christensen. 2002. Elk and land management. Pages 557-581 in: Toweill, D.E. and J.W. Thomas, Eds. North American Elk Ecology and Management. Smithsonian Institution Press, Washington, D.C.
McGarigal, K. 2002. Landscape pattern metrics. In: A. H. El-Shaarawi and W. W. Piegorsch, (eds.). Encyclopedia of Environmetrics Volume 2. John Wiley & Sons, Sussex, England. pp. 1135-1142.
McGarigal, K., S. A. Cushman, M. C. Neel, and E. Ene. 2002. FRAGSTATS: Spatial Pattern Analysis Program for Categorical Maps. Computer software program produced by the authors at the University of Massachusetts, Amherst. Available at the following web site:
www.umass.edu/landeco/research/fragstats/fragstats.html.
McGarigal, K., and B. J. Marks. 1995. FRAGSTATS: spatial pattern analysis program for quantifying landscape structure. USDA For. Serv. Gen. Tech. Rep. PNW-351.
Millington, A.C., X.M. Velez-Liendo, and A.V. Bradley. 2003. Scale dependence in multitemporal mapping of forest fragmentation in Bolivia: implications for explaining temporal trends in landscape ecology and applications to biodiversity conservation. ISPRS Journal of Photogrammetry and Remote Sensing 57: 289-299.
Moilanen, A., and M. Nieminen. 2002. Simple connectivity measures in spatial ecology. Ecology 83: 1131-1145.
Nagendra, H. and G. Utkarsh. 2003. Landscape ecological planning through multi-scale characterization of pattern: studies in the Western Ghats, south India. Environmental Monitoring and Assessment 87: 215-233.
Nepstad, D.C., A. Verissimo, A. Alencar, C. Nobre, E. Lima, P. Lefebvre, P. Schlesinger, C. Potter, P. Moutinho, E. Mendoza, M. Cochrane, and V. Brooks. 1999. Large-scale impoverishment of Amazonian forests by logging and fire. Nature 398: 505-508.
Newmark, W.D. 1995. Extinction of mammal populations in western North American national parks. Conservation Biology 9: 512-526. NGA (National Geospatial-Intelligence Agency). 2007. NGA Raster Roam, Vector Product Format Download (VMap Level 0). Available at:
http://geoengine.nga.mil/muse-cgi-bin/rast_roam.cgi
Noss, R.F., and A.Y. Cooperrider. 1994. Saving Nature’s Legacy. Island Press, Washington, DC. Phinney, J.T., F. Muller-Karger, P. Dustan, and J. Sobel. 2001. Using remote sensing to reassess the mass mortality of Diadema antillarum 1983-1994. Conservation Biology 15: 885-891.
Pickett, S.T.A. and P.S. White (eds.). 1985. The Ecology of Natural Disturbance and Patch Dynamics, Academic Press, 472 pages.
Oetter, D.R., Cohen ,W.B., Berterretche, M., Maiersperger, T.K. and R.E. Kennedy 2001. Land cover mapping in an agricultural setting using multiseasonal Thematic Mapper data. Remote Sensing of the Environment 76: 139-155.
O’Neill, R.V., K.H. Ritters, J.D. Wickham, and K.B. Jones. 1999. Landscape pattern metrics and regional assessment. Ecosystem Health 5(4): 225-233.
Reed, R.A., J. Johnson-Barnard, and W.L. Baker. 1996. Contribution of roads to forest fragmentation in the Rocky Mountains. Conservation Biology 10: 1098-1106.
Reice, S.R. 1994. Nonequilibrium determinants of biological community structure. American Scientist 82: 424-435.
Riitters K.H., , R.V. O’Neill, C.T. Hunsaker, J.D. Wickham. 1995. A factor analysis of landscape pattern and structure metrics, Landscape Ecology, 10 (1):23-39.
Riitters, K. H., Wickham, J. D., O’Neill, R., Jones, B., & Smith, E. (2000). Global-scale patterns of forest fragmentation. Conservation Ecology, 4(2), Art. 3.
Ritters, K.H., J.D. Wickham, and J.W. Coulston. 2004. A preliminary assessment of the Montreal Process indicators of forest fragmentation for the United States. Environmental Monitoring and Assessment 91: 257-276.
Rutledge, D. 2002. Landscape indices as measures of the effects of fragmentation: can pattern reflect process? Manuscript prepared for the New Zealand Department of Conservation. Science and Research Unit, Wellington, New Zealand.
Sanderson, E.W., M. Jaiteh, M.A. Levy, K.H. Redford, A.V. Wannebo, and G. Woolmer. 2002. The human footprint and the last of the wild. Bioscience 52: 891-904.
Schumaker, N.H. 1996. Using landscape indices to predict habitat connectivity. Ecology 77: 1210-1225.
Senay, G.B. and R.L. Elliot. 2002. Capability of AVHRR data in discriminating rangeland cover mixtures. International Journal of Remote Sensing 23(2):299- 312. doi:10.1080/01431160010014855
Sinclair, A.R., E.D.S. Hik, O.J. Schmitz, G.G.E. Scudder, D.H. Turpin, and N.C. Larter. 1995. Biodiversity and the need for habitat renewal. Ecological Applications 5(3): 579-587.
Soule, M.E., and J. Terborgh. 1999. Continental Conservation: Scientific Foundations of Reserve Networks. Island Press, Washington, DC.
Tischendorf, L. 2001. Can landscape indices predict ecological processes consistently? Landscape Ecology 16: 235-254.
Tischendorf, L. and L. Fahrig. 2000. On the usage and measurement of landscape connectivity. Oikos 90: 7-19.
Thomson, J.L., T. Schaub, N.W. Culver, and P. Aengst. 2005. Wildlife at a Crossroads: Energy Development in Western Wyoming. The Wilderness Society, Washington D.C.
Trombulak, S.C., and C.A. Frissell. 2000. Review of ecological effects of roads on terrestrial and aquatic communities. Conservation Biology 14: 18-30.
Turner, M.G., W.H.Romme, R.H. Gardner, R.V. O’Neill, and T.K. Kratz. 1993. A revised concept of landscape equilibrium: Disturbance and stability on scaled landscapes. Landscape Ecology 8: 213-227.
Vosselman, G. 1996. Uncertainty in GIS Supported Road Extraction, ISPRS Congress, Vienna, AIPRS, Vol.31, Part B3.
Vogt, P., Riitters, K.H., Estreguil, C., Kozak, J., Wade, T.G. and J.D. Wickham. 2007a. Mapping spatial patterns with morphological image processing. Landscape Ecology 22:171-177.
Vogt, P., Riitters, K.H., Iwanowski. M., Estreguil, C., Kozak, J., and P. Soille. 2007b. Mapping landscape corridors. Ecological Indicators 7:481-488.
Wade, T. G., Riitters, K. H., Wickham, J. D., & Jones, K. B. (2003). Distribution and Causes of Global Forest Fragmentation. Conservation Ecology, 7(2), art.7.
Weller, C., J. Thomson, P. Morton, and G. Aplet. 2002. Fragmenting Our Lands: The Ecological Footprint from Oil and Gas Development. The Wilderness Society, Washington, D.C.
Wickham, J.D., and K.H. Ritters. 1995. Sensitivity of landscape metrics to pixel size. International Journal of Remote Sensing 16: 3585-3594.
Wiens, J.A., C.S. Crawford, and J.R. Gosz. 1985. Boundary dynamics: A conceptual framework for studying landscape ecosystems. Oikos 45: 421-427.
Wilcove, D.S. (1989) Protecting biodiversity in multiple-use lands: lessons from the U.S. Forest Service. Trends in Ecological Evolution. 4: 385-388.
Wilcox, B.A., and D.D. Murphy. 1985. Conservation strategy: The effects of fragmentation on extinction. American Naturalist 125:879-887.
Wyoming Game and Fish Department (WGFD). 2004. Minimum Recommendations for Development of Oil and Gas Resources Within Crucial and Important Wildlife Habitats on BLM Lands. Wyoming Game and Fish Department, Cheyenne, WY.