Chapter 11. Trends in Invasive Alien Species
Authors:
Emma Underwood, Dept. of Environmental Science and Policy, University of California—Davis; IUCN Invasive Species Specialist Group
Susan Ustin, California Space Institute Center of Excellence, University of California—Davis
Contributors:
Aníbal Pauchard, Universidad de Concepción and Institute of Ecology and Biodiversity, Chile
Mathieu Maheu-Giroux, Universidad de Concepción and Institute of Ecology and Biodiversity, Chile
Reviewers:
Maj De Poorter, IUCN Invasive Species Specialist Group
Michael Browne, IUCN Invasive Species Specialist Group
Remote sensing based indicators for Invasive Alien Species
- Area, distribution and trends in particular invasive alien species
- Prediction of the distribution of invasive alien species
- Indirect identification of areas vulnerable to invasion
- Identification of potential sources of invasion and dispersal
11.1 Remote sensing technologies for mapping invasive alien species
Almost every ecosystem on Earth has serious problems with Invasive Alien Species (IAS), with invasions into natural systems representing a key threat to global biodiversity and ecosystem functioning as well as incurring economic costs (Mooney & Cleland 2001; Pimentel et al. 2005). Remote sensing and Geographic Information System (GIS) technologies offer potentially valuable tools for mapping and monitoring IAS as well as providing data inputs for predicting areas susceptible to invasion. Cost effective, large scale, and long term documentation and monitoring of IAS are recognized as fundamental research needs (Johnson 1999) which are increasingly being addressed. Over the last decade the number of publications on remote sensing applications to invasion biology has grown from 20 to 80 publications (Joshi et al. 2004).
The indicators in the beginning of this chapter outline a number of IAS indicators, both direct and indirect, across a range of scales that can be provided by remote sensing. The IAS problem and exchange of species between areas is by its nature global in extent, however, detection and invasion processes occur at more localized scales such as at the site or landscape scale, which is where remote sensing can contribute adequate mapping accuracies to be considered for operational use. Once IAS have become established in a region remote sensing, in comparison to field based techniques, allows an entire region to be mapped simultaneously, image-derived locations of IAS provide a permanent record that can be input into GIS databases for control activities, and repeated acquisitions allow trends in IAS abundance and distribution patterns to be efficiently monitored over time. In contrast, in areas where an IAS does not yet occur, prevention is the most efficient way of dealing with problematic species. Consequently, using remote sensing techniques to map points at risk is valuable, such as mapping airstrips in remote locations or seaports. Alternatively, once an IAS is established in a region mapping potential pathways with imagery can help prevent spread into new areas. For example, roads, hiking and horse trails, and off-road vehicle trails can be mapped—which disperse seeds on vehicle tires or on footwear. Similarly, boat launches can be identified since turbines on boat engines can spread fragments of aquatic invasive plants as well as small invasive aquatic animals such as snails. Since techniques for mapping these features are not specific to identifying IAS we do not review these approaches in this chapter.
Throughout this chapter, for both plants and animals, we use the term “Invasive Alien Species” (IAS). This replaces the various terminologies used in the studies reviewed which includes weeds, non-native, non-indigenous, exotic, alien, or introduced species (Richardson et al. 2000). We also focus primarily on detection of invasive plants or native plants impacted by an invasive species (e.g., pathogen), as these have received more attention using remote sensing applications than mapping invasive animals.
11.2 How does remote sensing distinguish invasive alien species?
The characteristics of a particular invasive plant species, both of the individual and also in relation to its surroundings, determine whether remote sensing techniques are appropriate. Many plant species, however, are not appropriate for image-based detection because they are indistinguishable from surrounding species or are understory species whose direct detection from remote sensing is almost impossible. In these cases indirect methods of mapping including the use of GIS data layers and modelling can be used (Joshi et al. 2004). For IAS present in the vegetation canopy successful detection approaches have generally capitalised on unique phenological or biochemical properties, structural characteristics, or the spatial patterns of infestations.
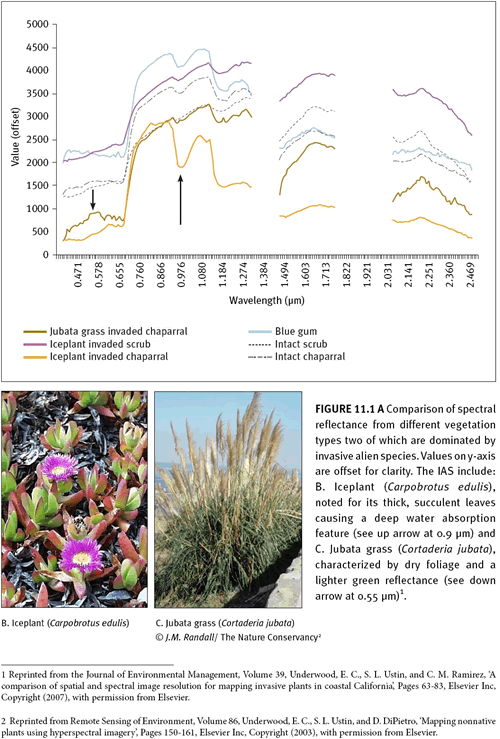
11.2.1 Biochemical characteristics Remote sensing images record the reflectance spectra of vegetation (and other land-cover elements such as soils and geologic minerals) within each pixel (picture element) based on their interactions with electromagnetic radiation in the solar region. Spectral characteristics of plants are derived from biochemical absorption features related to chlorophyll and other pigments, water, proteins, starches, waxes, and structural carbohydrate molecules such as lignin and cellulose (Elvidge 1990; Fuentes et al. 2001; Penuelas et al. 1997). Identification generally targets differences in the abundance and timing of these characteristics among species. For example, the strong water-absorption features associated with the succulent leaves of pickleweed (
Salicornia virginica) (Sanderson et al. 1998) or iceplant (
Carpobrotus edulis) (See figure 11.1) influence spectral properties which are detectable by remote sensors.
11.2.2 Phenological characteristics Detection by remote sensing can be facilitated by the timing of image acquisition to correspond with particular periods during the life cycle of a target IAS, or the acquisition of two images at different times in the life cycle. Some IAS become greener more quickly than surrounding species in the spring or alternatively senesce earlier, e.g., cheatgrass (
Bromus tectorum) or yellow starthistle (
Centaurea solstitialis) (Miao et al. 2006). Alternatively, the flower colour of the target invader may be distinct compared to surrounding species, such as the yellow bracts of leafy spurge (
Euphorbia esula).
11.2.3 Structural characteristics Where the canopy cover of the plant infestation is either dense or uniform the spatial patterning of the target species is often detectable. For example, the rhizomatous spread of iceplant (
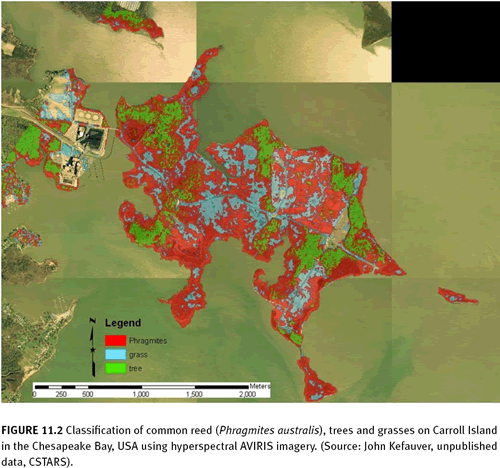
Carpobrotus edulis) produces a dense, uniform cover that assists in detection (See figure 11.1). Alternatively, detection can be assisted by the canopy architecture—branching patterns and leaf attachments—of IAS compared to the surrounding vegetation. For example, structural differences between common reed (
Phragmites australis) and the surrounding grass and tree vegetation types allowed for successful mapping of this invasive (See figure 11.2). Alternatively, the use of high spatial resolution lidar imagery which can detect small changes in the height of vegetation has been used in conjunction with aerial photography for mapping IAS in salt marshes and wetlands, e.g., Spartina spp. (Rosso et al. 2005).
11.3 Types of imagery for identifying invasive alien species
The application of remote sensing to invasive alien plants has seen a transition from the use of imagery with a high spatial but low spectral resolution (such as black and white or colour infrared aerial photographs) to digital images with greater spectral resolution although coarser spatial resolution, and more recently to imagery that couples both high spatial and high spectral resolution.
11.3.1 Aerial photography and videography At one end of the continuum of resolution options, aerial photographs have the benefit of being relatively inexpensive and typically have very fine spatial resolution (0.1–2 m). This spatial resolution meets most criteria for sampling invasive plant species of current management interest even in the smallest patch sizes, albeit not for many understory species or grasses in mixed grasslands which may not form distinct patches. Aerial photography (digital or film) and digital videography are particularly appropriate where an IAS has unique visual characteristics that readily distinguish it from the surrounding vegetation (see case study 11.1 and figure 11.3).

Chinese tamarisk (
Tamarix chinensis) has been identified with aerial photography using its unique orange-brown colour prior to leaf drop (Everitt et al. 1996). High resolution infrared aerial photography was used to map a number of woody invasives, including blackberry (
Rubus fruticosus), European olive (
Olea europaea), and
Pinus species in the Mount Lofty Ranges of South Australia (Crossman & Kochergen 2002). Visual and computer assisted interpretation of digital infrared images were also used to successfully map invading Acacia species from surrounding native vegetation and other IAS in the fynbos biome of South Africa (Stow et al. 2000). In the aquatic realm, expert interpretation of colour aerial photographs was successful in mapping submerged aquatic vegetation in the Chesapeake Bay, Maryland, USA (Orth & Moore 1983). While aerial photography is relatively inexpensive and can be acquired at high spatial resolution, disadvantages include extensive manual labor for processing and time-intensive interpretation requiring both skill and experience (Anderson et al. 1993; Everitt et al. 1995). Also, to capture spectral differences in the target species compared to surrounding vegetation image acquisition needs to be carefully coordinated with the timing of field measurements. Given these limitations data collection is feasible only over relatively small geographic areas.
Case study 11.1: Acacia dealbata invasion across multiple scales: Conspicuous flowering species can help us study invasion pattern and processes.
Authors: Aníbal Pauchard and Mathieu Maheu-Giroux
Multiscale approaches are powerful tools to understand the processes, patterns and impacts of biological invasions (Pauchard & Shea 2006).
While some IAS can be difficult to detect with remote sensing techniques, other species are particularly detectable due to their distinct spectral signature and unique phenology.
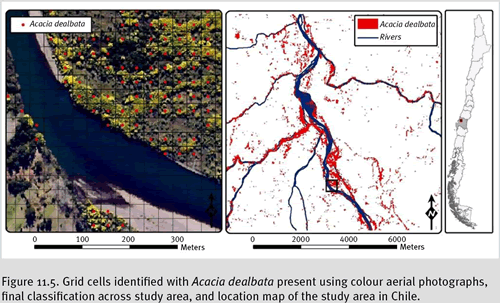
Acacia dealbata (silver wattle) is an invasive species in south-central Chile native to Australia, which flowers in the middle of winter, providing a clear and intense yellow pattern that can be distinguished using colour aerial photography (see figure 11.4).
 Acacia dealbata
Acacia dealbata is associated with human disturbance, particularly road construction, and invades many areas in the coastal range of south-central Chile as well as riparian corridors. Competitive effects with native forests are still unknown, as it is currently associated with disturbed environments such as exotic tree plantations (e.g.,
Pinus or
Eucalyptus). To determine the impact of
A. dealbata across multiple scales, we first assess the current extent of invasion across the landscape by taking advantage of the species’ winter yellow flower, which is unique compared to any other species in the region (native or exotic). We randomly selected three landscape quadrants (9 km x 9 km). In each quadrant, we georectified 1:20,000 digital colour photographs acquired in the winter using ortho-rectified images (1:115,000) and a minimum of 25 ground control points (using ArcGIS 9 software). The positional accuracy of the georectification was assessed using more than 30 reference points and the Root Mean Square Error was 8.45-13.65 m (Green & Hartley 2000). To maintain positional accuracy in our datasets, the minimum mapping unit needed to be at least twice the maximum RMSE (Ford & C.I. 1985; Walsh et al. 1987); consequently, a pixel size of 30 metres was chosen to record the presence/absence of
A. dealbata. A vector grid consisting of 30 m x 30 m cells was created and overlaid on each landscape quadrat. In each pixel, using visual photo-interpretation, presence of
A. dealbata was recorded when more than 5% of the pixel was occupied by the species (See figure 11.5).
Figure 11.5. Grid cells identified with
Acacia dealbata present using colour aerial photographs, final classification across study area, and location map of the study area in Chile.
To increase our understanding of the invasion of
A. dealbata, we analysed the relationship between the mapped infestations with a number of data layers (e.g., roads, rivers, land use). Preliminary results derived using logistic regression techniques show the species is significantly associated with rivers but less so with roads (p<0.01). There is significant aggregation of the species from the stand to the landscape scale (Modified Ripley’s K, L-function transformation (Besag 1977)), which illustrates that the species has a continuous distribution over the landscape once it establishes itself in a new location. This pattern may also indicate that the species is still spreading over the landscape and that limitations for establishment may be more related to propagule pressure than to environmental conditions.
Our current research focuses on relating species invasions to multiple environmental variables at different spatial scales and identifying how those relationships change as the scale of observation changes. For that purpose, we are considering a range of pixel size from 5 m to 150 m and also adjusting the size of the quadrants. For example, using the same aerial photographs, processes such as dispersal of the invasion front and percolation of the species into native ecosystems could be identified using 5 m pixels and quadrants of 1 km x 1 km.
Our methodological approach is relatively easy to conduct and replicate and requires neither highly skilled personnel nor expensive software nor hardware. However, due to the human-based decision process, biased and random errors can influence the presence/absence pixel classification. This can be especially problematic if the photographs are not acquired at peak flowering. This study indicates there is a tremendous potential for monitoring the spread of invasions of
A. dealbata in Chile using a relatively low-cost option of digital aerial photography, which can be readily applied to other IAS with distinct spectral characteristics. In turn, patterns identified using conspicuous flowering species can serve as models to increase our understanding of IAS, particularly of those that are difficult to detect using remote sensing techniques at broader spatial scales.
11.3.2 Multispectral imagery In contrast to aerial photography and videography, the use of digital multispectral imagery offers coverage over larger spatial areas, objective change detection through direct analysis of historical image archives, and the opportunity for automated image processing. Even coarse scale AVHRR imagery has been used to distinguish moderate to heavy infestations of broom snakeweed (
Gutierrezia sarothrae) from surrounding grassland species by capitalising on differences in phenological activity (Peters et al. 1992). Measurements of water clarity and turbidity from AVHRR images collected in different seasons were found to be accurate indicators of the locations of zebra mussels (
Dreissena polymorpha) in Lake Huron, USA (Budd et al. 2001). Clearly, these examples represent IAS that have spatially extensive distributions and background conditions that remain relatively consistent over large areas. At a medium spatial and spectral resolution, Landsat TM imagery has been used successfully to map target weed species that are spectrally or temporally unique. SPOT imagery has been used to successfully identify speargrass (
Imperata cylindrica) invading savanna areas in Cameroon (Thenkabail 1999). SPOT has also been suggested to be an appropriate sensor for monitoring the control of emergent aquatic invasive plants such as water hyacinth (
Eichhornia crassipies) in Bangalore, India using NDVI (Venugopal 1998). In one study, Landsat ETM+ imagery has been used to indirectly map IAS in the understory based on forest canopy density and light intensity reaching the understory in lowland forests in Nepal (Joshi et al. 2006). Multiband imagery offers some clear advantages over aerial photographs and videography but the coarse spatial and spectral resolution of AVHRR, MODIS, and Landsat (E)TM present challenges when infestations are neither continuously widespread, dense, nor monospecific. This might include some common and economically important invasive plants that mix with other species or have thin canopies that pass light from underlying vegetation and soil which can be surprisingly difficult to distinguish.
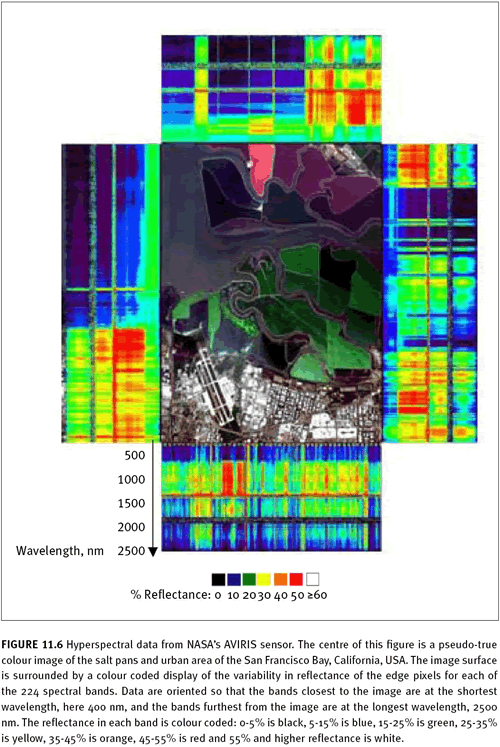
11.3.3 Hyperspectral imagery New technologies such as hyperspectral imagery (also known as imaging spectroscopy) hold great promise for mapping IAS. This technology, characterized by many narrow spectral bands, allows detailed spectra to be acquired for each pixel in an image (see figure 11.6). Subtle differences in reflection and absorption patterns can be detected resulting in the identification of individual species, higher mapping accuracies, and even the potential for mapping IAS that grow at low densities. As with other types of imagery, discrimination of the target species based on the distinctive colour of the target plant has been successfully used for mapping. For example, the white flowers of perennial pepperweed (
Lepidium latifolium) allowed this species to be discriminated in the Sacramento-San Joaquin Delta region of central California, USA (Andrew & Ustin 2006). In freshwater systems, again in the Delta, the greenness of water hyacinth (
Eichhornia crassipes) compared to surrounding vegetation permitted successful mapping of this emergent invasive (Mulitsch & Ustin 2003). In contrast to other image types, the increased spectral resolution of hyperspectral imagery is able to capitalise on specific biochemical and structural properties of target invaders, e.g., cellulose and lignin features caused by the dry foliage and stems of jubata grass (
Cortaderia jubata) (see figure 11.1C) and the nitrogen and water content of the fire tree (
Myrica faya) in Hawaii (Asner & Vitousek 2005; Underwood et al. 2007)
Hyperspectral imagery offers a number of advantages over coarser resolution sensors, although mapping IAS is still largely restricted to those which dominate the vegetation canopy. Disadvantages include the small number of airborne image providers, its expense, the large file sizes associated with the imagery and the expertise often required for processing.
11.4 Tradeoffs between image resolution and mapping accuracy
The selection of image resolution has clear implications for the accuracy of mapping target IAS or infested vegetation communities. However, while systematic comparisons of different types of imagery over the same spatial area would be valuable for comparison, there are few valid examples for IAS, and consequently limited guidelines to help inform the selection of appropriate image types. A study mapping invasive plants in riparian areas in South Africa found vegetation identification was most accurately derived with 1:10,000 black and white aerial photographs using manual techniques, while coarser spatial resolution aerial videography and Landsat TM imagery yielded lower accuracy (Rowlinson et al. 1999). Low mapping accuracies with the Landsat image were attributed to the large pixel size relative to the scale of riparian vegetation distribution. In Montreal, Canada, the accuracy of similar resolution panchromatic and colour aerial photographs were compared to map a common reed (
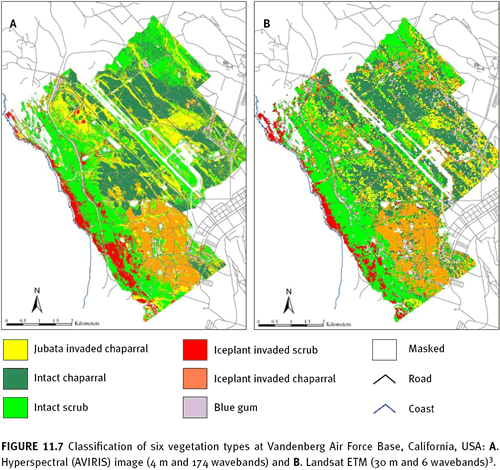
Phragmites australis) along roadsides and agricultural ditches (Maheu-Giroux & de Blois 2005). Findings indicated colour images with greater spectral resolution produced higher accuracy results. A study in the central coast of California, USA compared the accuracy of mapping six vegetation types—three of which were dominated by three IAS: iceplant (
Carpobrotus edulis), jubata grass (
Cortaderia jubata), and blue gum trees (
Eucalyptus globulus) (Underwood et al. 2007). Overall mapping accuracies ranged from 75% using hyperspectral (AVIRIS) imagery (4 m and 174 wavebands) to 50% using Landsat ETM imagery (30 m and 6 wavebands) (See figure 11.7). By comparing imagery with different combinations of spatial and spectral resolutions, findings suggest for IAS with distinct characteristics outside the visible spectrum that mapping accuracies are highest when imagery has greater spectral than spatial resolution.
Deciding on the appropriate image resolution is further confounded by the well recognized tradeoff between spectral and spatial resolution and the cost and ease of processing the data. Hunt et al. (2005) compared different types of imagery for a study area of 100 km2 and found Landsat TM to be cheapest at less than $500, followed by digital videography or moderate resolution imagery (e.g., SPOT) at less than $1,000, with hyperspectral imagery the most expensive at around $10,000 for the same size of study area. Again, as with mapping accuracy, there have been few standardized comparisons of the costs of remote sensing techniques over the same study area.
11.5 Identifying the potential distribution of invasive alien species
Although not a substitute for regular systematic field surveys, distribution models can provide a valuable tool to predict locations where undetected populations of particular species are most likely to occur. This can assist in prioritising limited resources such as personnel, time, and funding for control and monitoring purposes. Many predictive models consist of statistical “niche modelling”, where presences (e.g., from locations where herbarium specimens were collected) or presence-absence (e.g., from vegetation plots) data are used to infer an “environmental envelope” that contains most of the presence locations. Remotely sensed data such as vegetation, climate, soils, geomorphology, adjacency to roads or footpaths, and information on management techniques (e.g., grazing intensity maps) can be used as predictors in these models.
Alternatively, mechanistic models are more likely to take advantage of species tolerances (e.g., soil, moisture, elevation, and temperature extremes) established by experimental studies and use remotely sensed data to identify locations with high suitability with respect to multiple stressors or limiting resources. As illustrated in the studies reviewed in the following paragraphs, predictive modelling approaches have been applied at the global, national, landscape and site spatial scales (Morisette et al. 2006; Schnase 2003; Thuiller et al. 2005).
At the national scale, a habitat suitability map for salt cedar (
Tamarisk spp.) was created across the 48 continental US states with an accuracy of 90% (Morisette et al. 2006). This was created by integrating field data with land cover data (MODIS, 1 km), NDVI data (250 m), and an Enhanced Vegetation Index using logistic regression modelling. Also at the national scale, the potential ranges of major plant invaders in South Africa were assessed using climatic envelope models for 71 invasive plants utilising known presence data collected by gridcells (approximate 25 km x 25 km) across the country and variables such as growth days per year and mean annual precipitation (Rouget et al. 2004). At the landscape scale, in a retrospective study in New South Wales, Australia, the location of locust infestations (
Chortoicetes terminifera) was successfully determined using habitat type and soil type and condition derived from Landsat multispectral scanner (MSS) data, which provided a useful tool for predicting breeding sites in the future (Bryceson 1991). In freshwater systems, a study classified submerged and emergent vegetation from colour infrared aerial photography over 12 years and in conjunction with other data on bathymetry, herbicide application, nutrient levels and turbidity was able to relate the distribution of aquatic vegetation and environmental factors, producing an inexpensive tool for resource management (Welch et al. 1988). An alternative approach to species by species modelling is to identify hotspots of invasion for multiple species. A collaborative effort between NASA and the US Geological Survey is combining vegetation plot data with coarse scale remotely sensed information and geostatistical modelling to determine hotspots of invasive plant richness after a wildfire in New Mexico, USA (Schnase 2003).
11.6 Indirect identification of areas vulnerable to invasion
The approaches described in this section apply to IAS which occur in open areas and largely dominate the vegetation canopy. However, there are a number of other landscape and site features which can be detected by remote sensing which can assist in indicating the vulnerability of an area to invasion. Remote sensing can be used to identify areas of inappropriate grazing levels which can result in plant invasions through a combination of transporting propagules and micro-disturbance (although in some cases grazing also controls some IAS). In this case, imagery can be used to identify areas of reduced plant cover and soil and vegetation erosion. Second, detecting changes in the landscape over time can identify areas of disturbance which might be vulnerable to invasions. These include both natural disturbances such as landslides or the frequency or intensity of wildfire, or human-related such as trails, tracks and roads expanding into natural areas. Third, fragmentation of the natural habitat results in edge habitats being more vulnerable to invasion by plants and animals than core habitats (Hobbs 1998). The spatial distribution of natural habitats in the imagery and fragmentation indices can help identify vulnerable areas.
11.7 Limitations of remote sensing and modelling applications to invasive alien species
11.7.1 Limitations for Remote Sensing Detection Utilising remote sensing to identify invasive plant species suffers from many of the same caveats as using imagery to map other land cover features. First, the cost of data as well as the necessary software and hardware to support them is high, particularly for the newer forms of high spatial and spectral resolution imagery, although the overall trend is towards declining imagery costs (Turner et al. 2003). Second, the technical expertise required for processing both aerial photography (expert interpretation) and hyperspectral imagery (image processing) are high, with processing techniques for hyperspectral data still in the research and development phase. Third, the ability to detect IAS and the accuracy with which detection can be achieved varies among ecosystems, for example, identifying invasive plants in freshwater systems is challenging compared to terrestrial systems since submerged species are difficult to distinguish from water when present at low density and water turbidity from sediment and/or algae can mask detection (Underwood et al. 2006). Finally, while remote sensing techniques permit greater efficiency, they can never entirely replace in situ field measurements. The best results generally occur when both field data and imagery are used together with field measurements providing critical inputs for classifying and validating image classifications. Analyses are also significantly improved by a realistic understanding of, and familiarity with, the habitat particularly because of changes that can occur with the plant life cycle and the implications of these on spectral properties.
Despite these limitations, remote sensing is the only method for efficiently collecting information over large spatial extents at high spatial resolution and with 100% spatial sampling. In the future, developments in remote sensing data such as the increasing temporal frequency of image acquisition, techniques which fuse both passive and radar sensors, and the use of thermal image and texture analysis are likely to increase the tools available for mapping IAS. Furthermore, the synoptic view of imagery means lands in both public and private land ownership can be examined and analysed as a continuous landscape, thus facilitating an understanding of how land use may interact with the spread of invasives. Tradeoffs between image resolution and costs are rapidly changing as these technologies mature and more data providers are available. Policies governing access to original data also continue to evolve, for example, in the distribution of US governmental satellite data the benefits of increased imagery requires organized, searchable, and well-documented libraries of processed imagery and models.
11.7.2 Limitations for Predictive Models Predictive models are valuable tools to inform decision making, however, there are a number of limitations. One of the obvious concerns regarding the output of species models is that predictions might over- or under-estimate the potential distribution of the target IAS. Over-prediction of distributions may result from identifying potential niche areas to which species may be unable to disperse (Peterson & Vieglais 2001), while under-prediction might result from insufficient explanatory variables used to correlate with the species occurrence. Furthermore, other data relevant for determining the spread of IAS—such as propagule pressure which determines the ease with which invasive plants overcome environmental barriers to become established (Rouget & Richardson 2003)—are generally not included. Second, predictive models are often static and fail to incorporate ecological processes and disturbances that might facilitate invasion, e.g., fire history or flood regimes. Finally, where known presence data are used, this is often based on a limited number of field collected data points within a discretely defined area, which is often at a finer spatial resolution than the selected explanatory data layers. To model at national or global spatial scales, occurrence data of the species across the range of explanatory variables are required.
Nonetheless, future research should emphasize the development of predictive models that incorporate remote sensing, environmental data and GIS techniques as a time and cost efficient method of identifying areas vulnerable to invasion. These approaches are particularly valuable for land managers with limited resources for monitoring and controlling IAS. Alternatively, predictive models can be used in aiding prevention of invasives by prioritizing species for which in-depth risk analyses should be undertaken. In both cases, results from models can aid the development of proactive policies at the national scale to restrict importation of high-risk IAS.
11.8 Data and other resources
The Global Invasive Species Programme (GISP) Aims include raising global awareness of the ecological and socio-economic impacts of IAS by developing a global information system on IAS, disseminating information on the impacts of invasives, providing information and training, and building international networks to achieve this.
The Global Invasive Species Database (GISD) The GISD was developed as part of the global initiative on IAS led by the Global Invasive Alien Species Programme. It aims to increase public awareness about IAS to facilitate effective management and contains information on the ecology, impacts, distribution and pathways of IAS.
The Global Invasive Species Information Network (GISIN) GISIN was formed to provide a platform for sharing IAS information at a global level, via the Internet and other digital means. Activities include developing a pilot system to search across diverse IAS information systems that are already present on the Internet.
A Weed Managers Guide to Remote Sensing and GIS The website provides technical information and guidance to help resource managers learn to use remote sensing and GIS to map, monitor and predict invasions.
The Nature Conservancy: Global Invasive Species Network The website reviews remote sensing and GIS technologies for detecting invasive species, particularly in their natural environment.
Weed Invasion Susceptibility Prediction The website provides an extension that can be used with GIS software (ArcView) to predict areas susceptible to invasion by IAS and also their predicted spread over time.
Invasive Species Forecasting System
NASA and US Geological Survey are developing a forecasting system for the early detection, remediation, management, and control of invasive species
Acknowledgements
We wish to acknowledge sponsors of several projects of the
Center for Spatial Technologies and Remote Sensing (CSTARS) which are cited in this chapter including the
Strategic Environmental Research and Development Program (SERDP) joint DOD/EPA/DOE conservation program for grant “Application of hyperspectral techniques to monitoring and management of invasive weed infestation”, the NSF Complex Systems program “Dynamics of an invasive non-native species. Predicting and understanding spread and its impacts, biological, physical and human: the case of Spartina alternaflora in the western US”, and the NASA/EPA STAR program for biological indicators of wetland ecosystems health, the “Western Center for Estuarine Ecosystem Indicator Research”, and the NSF IGERT Training grant “Biological Invasions: From genes to ecosystems, from science to society”, and lastly the NASA Airborne Science program at the Jet Propulsion Laboratory for support in collection of AVIRIS datasets and the HyVista Corporation for HyMap data used in this chapter.
11.9 References
Anderson, G. L., J. H. Everitt, A. J. Richardson, and D. E. Escobar. 1993. Using satellite data to map false broomweed (
Ericameria austrotexana) infestations on south Texas rangelands. Weed Technology 7:865– 871.
Andrew, M. E., and S. L. Ustin. 2006. Spectral and physiological uniqueness of perennial pepperweed (
Lepidium latifolium). Weed Science 54:1051-1062.
Asner, G. P., and P. M. Vitousek. 2005. Remote analysis of biological invasion and biogeochemical change. Proceedings of the National Academy of Sciences of the United States of America 102:4383-4386.
Besag, J. 1977. Some methods of statistical analysis for spatial data. Bulletin of the International Statistical Institute 47:77-92.
Bryceson, K. P. 1991. Likely locust infestation areas in western New South Wales, Australia, located by satellite. Geocarto International 6:21-37.
Budd, J. W., T. D. Drummer, T. F. Nalepa, and G. K. Fahnenstiel. 2001. Remote sensing of biotic effects: zebra mussels (
Dreissena polymorpha) influence on water clarity in
Saginaw Bay, Lake Huron. Limnology and Oceanography 46:213-223.
Crossman, N. D., and J. Kochergen. 2002. Mapping environmental weeds in the Mount Lofty Ranges, South Australia, using high resolution infrared aerial photography. Pages 1-4 in H. Spafford Jacob, J. Dodd, and J. H. Moore, editors. Proceedings of the thirteenth Australian weeds conference. Plant Protection Society of Western Australia, Perth, Australia.
Elvidge, C. D. 1990. Visible and near infrared reflectance characteristics of dry plant materials. International Journal of Remote Sensing of Environment 11:1775–1795.
Everitt, J. H., G. L. Anderson, D. E. Escobar, M. R. S. Davis, N. R., and R. J. Andrascik. 1995. Use of remote sensing for detecting and mapping leafy spurge (
Euphorbia esula). Weed Technology 9:599-609.
Everitt, J. H., D. E. Escobar, M. A. Alaniz, M. R. Davis, and J. V. Richerson. 1996. Using spatial information technologies to map Chinese tamarisk (
Tamarix chinensis) infestations. Weed Science 44:194-201.
Ford, G. E., and C.I. Zanelli. 1985. Analysis and quantification of errors in the geometric correction of satellite images. Photogrammetric Engineering & Remote Sensing 51.
Fuentes, D. A., J. A. Gamon, H. L. Qiu, D. A. Sims, and D. A. Roberts. 2001. Mapping Canadian boreal forest vegetation using pigment and water absorption features derived from the AVIRIS sensor. Journal of Geophysical Research 106:33565-33577.
Green, D. R., and S. Hartley. 2000. Integrating photointerpretation and GIS for vegetation mapping: some issue of error. Pages 103-134 in R. Alexander, and A. C. Millington, editors. Vegetation Mapping: From Patch to Planet. John Wiley & Sons Ltd, Chichester, U.K.
Hobbs, R. J. 1998. Impacts of land use on biodiversity in southwestern Australia. Pages 81-106 in P. W. Rundel, G. Montenegro, and F. M. Jaksic, editors. Landscape Disturbance and Biodiversity in Mediterranean-Type Ecosystems. Springer-Verlag, Berlin.
Hunt, R., R. Hamilton, and J. Everitt. 2005. A weed manager’s guide to remote sensing and GIS (rsac-0077). Mapping weed infestations using remote sensing
http://www.fs.fed.us/eng/rsac/invasivespecies/documents/mapping.pdf Pages 1-8.
Johnson, D. E. 1999. Surveying, mapping, and monitoring noxious weeds on rangelands. Pages 19-35 in R. L. Sheley, and J. K. Petroff, editors. Biology and management of noxious rangeland weeds. Oregon State University Press, Corvallis, Oregon.
Joshi, C., J. de Leeuw, J. van Andel, A. K. Skidmore, H. D. Lekhak, I. C. van Duren, and N. Norbu. 2006. Indirect remote sensing of a cryptic forest understory invasive species. Forest Ecology and Management 225:245-256.
Joshi, C., J. de Leeuw, and I. C. van Duren. 2004. Remote sensing and GIS applications for mapping and spatial modelling of invasive species. Pages 669-677. Proceedings for ISPRS, Istanbul, Turkey.
Maheu-Giroux, M., and S. de Blois. 2005. Mapping the invasive species
Phragmites australis in linear wetland corridors. Aquatic Botany 83:310-320.
Miao, X., P. Gong, S. Swope, R. L. Pu, R. Carruthers, G. L. Anderson, J. S. Heaton, and C. R. Tracy. 2006. Estimation of yellow starthistle abundance through CASI-2 hyperspectral imagery using linear spectral mixture methods. Remote Sensing of Environment 101:329-341.
Mooney, H. A., and E. E. Cleland. 2001. The evolutionary impact of invasive species. Proceedings of the National Academy of Sciences 98:5446-5451.
Morisette, J. T., C. S. Jarnevich, A. Ullah, J. Cai W, J. A. Pedelty, J. E. Gentle, T. J. Stohlgren, and J. L. Schnase. 2006. A tamarisk habitat suitability map for the continental United States. Frontiers in Ecology and the Environment 4:11-17.
Mulitsch, M., and S. L. Ustin. 2003. Mapping invasive plant species in the Sacramento-San Joaquin Delta region using hyperspectral imagery. Report to the California
Department of Boating and Waterways. Center for Spatial Technologies and Remote Sensing, University of California, Davis, Sacramento, California.
Orth, R. J., and K. A. Moore. 1983. Submerged vascular plants: techniques for analysing their distribution and abundance. Marine Technology Society Journal 17:38-52.
Pauchard, A., and K. Shea. 2006. Integrating the study of non-native plant invasions across spatial scales. Biological Invasions 8:399-413.
Penuelas, J., J. Pinol, R. Ogaya, and I. Filella. 1997. Estimation of plant water concentration by the reflectance Water Index WI (R900/R970). International Journal of Remote Sensing 18:2869-2875.
Peters, A. J., B. C. Reed, M. D. Eve, and K. C. McDaniel. 1992. Remote sensing of broom snakeweed (
Gutierrezia sarothrae) with NOAA-10 spectral image processing. Weed Technology 6:1015–1020.
Peterson, A. T., and D. A. Vieglais. 2001. Predicting species invasions using ecological niche modeling: new approaches from bioinformatics attack a pressing problem. BioScience 51:363-371.
Pimentel, D., R. Zuniga, and D. Morrison. 2005. Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecological Economics 52:273-288.
Richardson, D. M., P. Pysek, M. Remanek, M. G. Barbour, F. D. Panetta, and C. J. West. 2000. Naturalization and invasion of alien plants: concepts and definitions. Diversity and Distributions 6:93-107.
Rosso, P. H., S. L. Ustin, and A. Hastings. 2005. Use of lidar to study changes associated with Spartina invasion in San Francisco Bay marshes. Remote Sensing of Environment 100:295-306.
Rouget, M., and D. M. Richardson. 2003. Inferring process from pattern in plant invasions: A semimechanistic model incorporating propagule pressure and environmental factors. American Naturalist 162:713-724.
Rouget, M., D. M. Richardson, J. L. Nel, D. C. Le Maitre, B. Egoh, and T. Mgidi. 2004. Mapping the potential ranges of major plant invaders in South Africa, Lesotho and Swaziland using climatic suitability. Diversity and Distributions 10:475-484.
Rowlinson, L. C., M. Summerton, and F. Ahmed. 1999. Comparison of remote sensing data sources and techniques for identifying and classifying alien invasive vegetation in riparian zones. Water SA 25:497-500.
Sanderson, E. W., M. Zhang, S. L. Ustin, and E. Rejmankova. 1998. Geostatistical scaling of canopy water content in a California salt marsh. Landscape Ecology 13:79–92.
Schnase, J. L. 2003. Invasive species forecasting system. Test site – Cerro Grande Wildfire Site.
http://invasivespecies.gsfc.nasa.gov/isfs_cerro.html. NASA/GSFC.
Stow, D., A. Hope, D. Richardson, D. Chen, C. Garrison, and D. Service. 2000. Potential of colour-infrared digital camera imagery for inventory and mapping of alien plant invasions in South African shrublands. International Journal of Remote Sensing 21:2965 – 2970.
Thenkabail, P. S. 1999. Characterization of the alternative to slash-and-burn benchmark research area representing the Congolese rainforests of Africa using near-real-time SPOT HRV data. International Journal of Remote Sensing 20:839-877.
Thuiller, W., D. M. Richardson, P. Pysek, G. F. Midgley, G. O. Hughes, and M. Rouget. 2005. Niche-based modelling as a tool for predicting the risk of alien plant invasions at a global scale. Global Change Biology 11:2234-2250.
Turner, W., S. Spector, N. Gardiner, M. Fladeland, E. Sterling, and M. Steininger. 2003. Remote sensing for biodiversity science and conservation. Trends In Ecology & Evolution 18:306-314.
Underwood, E. C., M. J. Mulitsch, J. A. Greenberg, M. L. Whiting, S. L. Ustin, and S. C. Kefauver. 2006. Mapping invasive aquatic vegetation in the Sacramento-San Joaquin Delta using hyperspectral imagery. Environmental Monitoring and Assessment 121:47-64.
Underwood, E. C., S. L. Ustin, and C. M. Ramirez. 2007. A comparison of spatial and spectral image resolution for mapping invasive plants in coastal California. Journal of Environmental Management 39:63-83.
Venugopal, G. 1998. Monitoring the effects of biological control of water hyacinths using remotely sensed data: a case study of Bangalore, India. Singapore Journal of Tropical Geography 19:92-105.
Walsh, S. J., D. R. Lightfoot, and D. R. Butler. 1987. Recognition and assessment of error in Geographic Information System. Photogrammetric Engineering and Remote Sensing 53:1423-1430.
Welch, R., M. Remillard, and R. B. Slack. 1988. Remote sensing and geographic information system techniques for aquatic resource evaluation. Photogrammetric Engineering & Remote Sensing 54:177-185.