Chapter 8. Trends in Species Populations
Authors:
Colby Loucks, World Wildlife Fund (WWF-US)
Peter Leimgruber, Smithsonian National Zoological Park
Contributors:
James Strittholt, Conservation Biology Institute
Hong Jiang, Conservation Biology Institute
Reviewers:
Sarat Babu Gidda, Secretariat of the Convention on Biological Diversity
Remote sensing based indicators for species populations:
- Location and extent of species aggregations
- Extent of specific habitats—direct detection
- Extent of specific habitats—indirect detection
- Extent of specific habitats—modelled
8.1 Background
Promoting the conservation of species diversity is one of the goals of the Convention on Biological Diversity’s 2010 target. To effectively attain this goal and its sub-targets would require the maintenance of viable populations of selected species. In turn, this requires a sufficient number of individuals, populations (different groups of individuals occupying the same area), and habitat (the space and biophysical features required to support the species). Therefore, basic measures of species diversity include number (species richness), abundance (population size) of species, number of viable populations, and habitat area.
Remote sensing data have become vital to monitoring trends in the distribution and abundance of many plant and animal species. Two general approaches are used. The first is to directly monitor individual organisms or populations, using airborne or satellite sensors. The indirect approach is to use remotely sensed environmental data in predictive models, estimating species location within a geographic information system (GIS) or statistical package. Although direct species monitoring via remote sensing (i.e. relating spectral reflectance data to species or biodiversity distribution) is relatively uncommon, the use of distribution prediction and modelling continues to increase as new remote sensing data, modelling tools, and statistical techniques develop and mature.
8.2 Remote sensing to directly monitor species populations
Currently, population trends of only a few relatively large or very abundant plant and animal species can directly be monitored using remote sensing techniques. McGraw et al. (1998) and Key et al. (2001) demonstrated success in predicting deciduous tree species distributions, though for relatively small areas. It is possible to use satellite imagery to map the extent and change of communities of colonial organisms such as coral reefs (Mumby et al. 1997; Hochberg and Atkinson 2000, also see Chapter 7), or even individual colonies of burrowing mammals such as prairie dogs (Sidle et al. 2001). However, most direct monitoring has focused on medium to large mammals or flocks of migratory birds in open landscapes (i.e., at stopover sites or in open wetlands, grasslands, or savanna). The latter involve airplane or helicopter aerial surveys, high resolution satellite data, or a combination. Some examples of direct monitoring include white-backed vulture (Murn et al. 2002), saiga antelope (Milner-Gulland et al. 2001), whales (Best 1990), elephants (Whitehouse et al. 2001), orangutan nests (Ancrenaz et al. 2005), and Australian mammals (Tracey et al. 2005).
The ability to directly monitor species is rare, however. The majority of species cannot be identified, counted, mapped, or monitored directly from satellite or aerial imagery for several reasons, including the size of the species in relation to sensor resolution, natural history (i.e., nocturnal species would be difficult to detect using optical sensors), and the “long return time” (i.e., the time it takes the satellite to return to the same spot on the earth for a second image). Even when detection of animals via satellite or airborne techniques is possible, there are still problems in determining whether the samples are statistically valid. Because of the number of images needed for an accurate estimate, direct monitoring will be prohibitively expensive in the near future for most candidate species.
8.3 Remote sensing to indirectly monitor species populations: modelling species distributions by mapping habit
Meanwhile, examples of indirect estimates of populations and distributions derived from remotely sensed data are increasing (Turner et al. 2003). Biophysical or other environmental parameters that can be measured are used to predict the distribution of habitats and species (Kerr and Ostrovsky, 2003). This approach combines knowledge of species distributions from on-the-ground field surveys with environmental parameters (e.g., habitat, elevation, rainfall) to develop an empirical model that can be applied over a much greater area (Manly et al. 2004)
The assumption that habitat area is an indicator of species richness is based on the species area relationship S = cAz, where S is the number of species, A is area, and c and z are constants (Arrhenius, 1921). Assumption of this equation allows habitat classifications to infer species losses associated with changes in land cover and land use. Where local parameters (c and z) of the species-area curve have been tested, clarifying the precise relationship between species richness and habitat area, remote sensing can complete the area component of the equation (Turner et al. 2003).
Threat mapping is an indirect way of mapping a species’ distribution or more precisely of mapping where species are not found. In contrast to species-habitat models that delineate areas potentially occupied by a given species, threat mapping may delineate areas that are avoided by a species. For example, Asian elephants (
Elephas maximus) tend to be habitat generalists; they can be found in most natural environments ranging from grasslands and wetlands to dense moist forests. However, in areas of dense agriculture and human population, they have a much greater risk of being killed as the result of crop raiding and human-elephant conflict. This type of “exclusion” or threat mapping can complement habitat mapping or be used by itself. Measured trends in habitat or threat extent can be used as a proxy for trends in the species’ population. Remote sensing and remotely sensed imagery are essential for this type of indirect monitoring of species.
A majority of the species distribution models correlate potential habitat or ecosystem properties (i.e., net primary productivity) with on-the-ground surveys to monitor population trends. However, the accuracy for mapping a specific habitat type is frequently between 75 and 85 percent. Therefore, we often cannot detect minor changes very accurately. Increasingly, scientists are attempting to map and delineate either biodiversity measures or species distribution directly from spectral reflectance information (such as Normalized Difference Vegetation Index contained in satellite images (Leimgruber et al. 2005). Using NDVI, Gould (2000) estimated species richness of three vascular species in Central Canadian Arctic region. The advantage of directly relating species distribution to an environmental variable is that there is no additional error introduced through the habitat classification process.
Habitat loss is the single most important factor in species extinction (Baillie et al. 2004). Consequently, global, continental, or countrywide habitat change mapping can be an important tool in the monitoring of remaining wild populations of species. Although it should not replace systematic on-the-ground surveys and monitoring for species populations, satellite data can and should play a significant role in the following:
- establishing baseline datasets on current extent of suitable habitat for a given species;
- identifying zones for on-the-ground survey/monitoring (for example, satellite data can be used to identify areas of habitat similar to those known to be occupied by a species, or areas where species are threatened by habitat loss);
- providing early warning systems for habitat loss and potential population decline; and
- determining fragmentation and connectivity of remaining populations and metapopulations.
8.4 Practical examples
Systematic monitoring of species populations at global to continental scales exists, but most studies have been done at the national scale or smaller. Examples of global and continental monitoring of mammals include the North American Breeding Bird Surveys (Sauer et al. 2005), monitoring of African elephants by the African Elephant Specialist Group,
Monitoring the Illegal Killing of Elephants (MIKE), and the
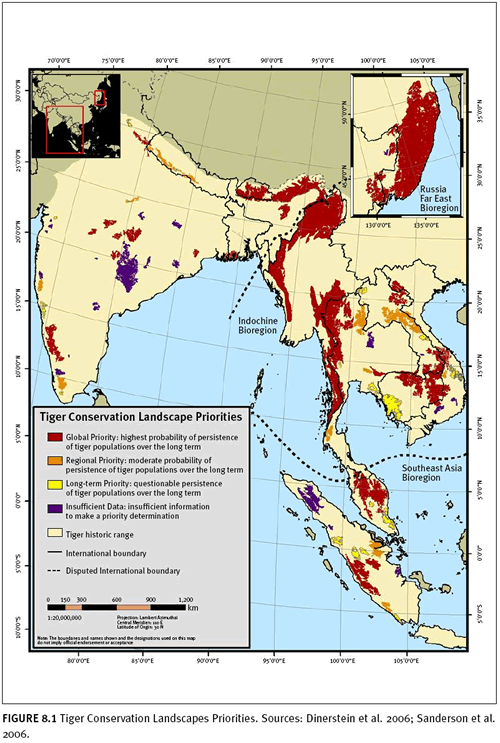
Save the Tiger Fund (STF) has spearheaded an effort to identify and prioritize the remaining habitat – Tiger Conservation Landscapes (TCLs) – for tigers across their range.
Countrywide or smaller-scale species distribution models are abundant, and they are increasingly using remote sensing data. Rushton et al. (2004) analysed 21 species distribution modelling studies published in the Journal of Applied Ecology since 2000. Of these studies, 16 used remote sensing or mapped habitat data, often in combination with field survey data, to predict species distribution for mammals, birds, lepidoptera, and reptiles. Recent studies include the following:
- Raxworthy et al. (2003) used old locality data from museums in combination with satellite data to predict the distribution of 11 chameleon species in Madagascar. They achieved an overall accuracy rate of 82 percent and also identified areas of “overprediction,” where the model suggested that there were more species than had been recorded. Once these areas were surveyed, they yielded seven chameleon species new to science.
- Loucks et al. (2003) used SPOT imagery to identify forest-nonforest habitat to identify potential giant panda (Ailuropoda melanoleuca) habitat in the Qinling Mountains of China. They combined the forest data with elevation and field data to develop a predictive model of habitat distribution and giant panda abundance. This analysis eventually led to the creation of six nature reserves to protect gaps in the giant panda’s habitat.
- McShea et al. (1999) used satellite maps of Eld’s deer (Cervus eldii) habitat in Myanmar to identify remaining habitat for this highly endangered species.
- Koy et al. (2005) expanded on this study and used a continuous-field analysis of Landsat data to predict tree density in open-canopy dipterocarp forests, the preferred habitat by Eld’s deer. They found that Eld’s deer distribution was related to tree density.
- Osborne et al. (2001) used Advanced Very High Resolution radiometer (AVHRR) imagery in conjunction with disturbance data to predict the distribution of great bustards in central Spain. They found that sites occupied by bustards had significantly lower densities of human-altered land (e.g., roads, buildings) and also occurred in a narrow range of elevations.
- Buchanan et al. (2005) successfully combined field survey data with a supervised classification of a Landsat image to identify detailed categories of moorland vegetation. They then were able to predict golden plover (Pluvialis apricaria) abundance, based on habitat associations, across large moorland areas.
Case Study 8.1: Mapping Habitat for the Giant Panda in Sichuan Province, China
Authors: Jim Strittholt and Hong Jiang (Conservation Biology Institute)
Indicators: forest cover, fragmentation, threatened species
Potential monitoring scales: regional, national
Sensor: Landsat Thematic Mapper (TM) and Enhanced Thematic Mapper (ETM+)
Imagery cost/hectare: free
Total project cost: $75,000
Limitations on accuracy: terrain shadows, cloud cover, important understory vegetation component
a. Introduction Sichuan Province is located in southwest China and is recognized as one of the world’s biodiversity hotspots. A wide range of natural environments occurs in the region, from alpine tundra to subtropical forests. The region supports high levels of species richness and centers of high species endemism. It is home to several of the world’s best-known mammals, including the giant panda (
Ailuropoda melanoleuca). The Sichuan Province also supports a growing human population that currently numbers nearly 90 million people. The intersection of globally significant biological values and high human pressures highlights this region as one of tremendous conservation importance, with considerable challenges. In response, the Chinese government has invested heavily over recent years to try to prevent the continuing degradation of natural ecosystems throughout the region, with special emphasis being paid to the giant panda—the symbol of the nation. Presently, there are 131 nature reserves, covering approximately 7,620,000 hectares (about 15.6 percent of the province), managed for conservation purposes to varying degrees. The purpose of this case study was to examine land cover change, fragmentation, and status of the giant panda for a large portion of its current range.
b. Methods We used remote sensing (Landsat TM 1986 and Landsat ETM+ 2002) to examine the major land cover changes, which we supplemented with a couple of important ancillary data layers (i.e., roads and bamboo distribution), as well as panda survey data for two dates: 1988 and 2004. We processed Landsat 5 TM imagery (circa 1986) and Landsat 7 ETM+ imagery (circa 2002), using ERDAS Imagine software, and mapped general land cover types with special attention to native forests (the prime habitat for giant pandas), using an unsupervised classification approach we developed: the Optimal Iterative Unsupervised Classification (OIUC) method (Jiang et al. 2004). For approximately 30 percent of the study area, we collected more than 700 ground control points, using the global positioning system (GPS). The accuracy of the classification was assessed by field validation and a previously published ancillary spatial database.
c. Results Results varied across the study area. For the northern portion, where commercial logging was halted in 1998, the loss of forest cover between the two sampling dates was significant, but less severe than in the southern portion, where commercial logging has continued at a rapid rate (figure 8.2). Liu et al. (2001) found similar results for the nearby Wolong Nature Reserve. Overall, image classification accuracy was estimated to be 92 percent for this region. The classification was correct 89 percent of the time for forests, 85 percent for shrublands, 92 percent for orchards, 93 percent for grasslands, 90 percent for croplands, and 98 percent for water. Bamboo extent and spatial distribution were also variable. In the north, the natural life cycle of maturity, flowering, decline, and regeneration were the driving force for the observed changes. In the south, loss of bamboo was noted primarily because of human activity. Habitat fragmentation via roads and agriculture continues throughout the study area. Panda survey points for both dates were closely associated with bamboo cover. Overall, the newest panda survey shows greater numbers of pandas in 2004 than in the previous 1988 survey. There may be several explanations, including that greater effort was spent on the latest survey than in previous attempts.
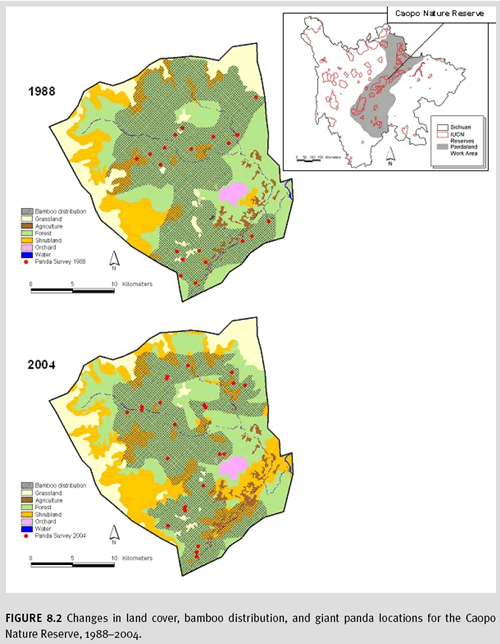
For the Caopo Nature Reserve, difference in the number of pandas sampled between the two dates was not statistically significant, but there were some notable changes in the physical locations being used by the animals. For example, some regions have lost pandas (e.g., around the agricultural lands) between the two survey dates, which may coincide with either the loss of forest habitat around these agricultural lands or from direct disturbance by associated human presence and activity. It remains to be seen whether these lands will become unsuitable giant panda habitat in the future and to what degree habitat fragmentation will become a limiting factor, especially given that pandas are so closely linked with bamboo, which is so naturally dynamic in its distribution.
d. Limitations The most important limitations in classifying satellite imagery for the mountains of southwest China include high levels of terrain shadows, cloud cover, and physical access for assessing accuracy. For our case study, the importance of mapping bamboo (an understory plant not easily discernable via remote sensing) was critical. Linderman et al. (2004) combined Landsat TM imagery and a nonlinear artificial neural network algorithm for mapping understory bamboo in the Wolong Nature Reserve and reported 80 percent accuracy. However, because we had to treat such a large area and field-based bamboo distribution maps were already available (including life-history stages: young, mature, flowering) for each of the survey years, we elected to convert the existing bamboo distribution maps (including life-history stages) for the two dates to computer-readable form by digitizing field maps. We were also very fortunate to acquire the national panda survey data results from 1988 and 2004 for all of Sichuan Province. Combining the necessary spatially explicit data on the changes in (1) forest habitat, (2) bamboo distribution, and (3) panda locations was extremely important for effective monitoring. Having this level of data for other species of concern around the world may be more challenging, but the importance of these multi-source data is an important finding.
8.5 Caveats
Inferring the number or distribution of species using a proxy (such as amount and configuration of habitat or primary productivity) assumes prior knowledge of species habitat preference. This information should be known with some degree of confidence before establishing a monitoring programme.
Even with complete knowledge of habitat preference, there is potential for over- or under-predicting the distribution of a species. For example, overestimating species range may occur if the remote sensing analysis predicts sufficient habitat for a given species, but in reality the species of interest is absent or limited for some other reason: perhaps lack of prey, disease, or poaching. The existence of multiple limiting factors, many of which cannot be remotely sensed, underscores the need to combine remote sensing analyses with on-the-ground surveys and local field knowledge.
Underestimation may occur when the remote sensing analysis either misclassifies the habitat or predicts the loss of habitat and therefore extirpation (local extinction) of a species—when the species is still present in reality. The likely cause of this error is that the definition of “habitat” for a species is too narrow, and that particular species is able to survive in a broader spectrum of habitats than originally known. Using a multistage sampling approach (i.e., employing remote sensing in conjunction with ground sampling) will help prevent this type of error. Again, the use of local knowledge—both when conducting ground surveys and when classifying imagery—is invaluable in minimizing this type of error.
Despite these shortcomings, remotely sensed mapping of habitat often provides a low-cost first approximation of the population status for many species. In addition, it can provide valuable information on how and where to survey for remaining populations of species. It represents a very powerful monitoring tool when employed together with finer-scale monitoring and field analysis.
8.6 Resources
The following are examples of software specifically made for habitat modelling. Models can be constructed within a simple GIS as well.
MaxEnt:
http://www.cs.princeton.edu/~schapire/maxent/ DivaGIS:
http://research.cip.cgiar.org/confluence/display/divagis/Home;jsessionid=24956A8AE866D4124661AA06872D8B5B GARP:
http://www.nhm.ku.edu/desktopgarp/ Biomapper:
http://www2.unil.ch/biomapper/
8.7 References
Ancrenaz M., Gimenez O., Ambu L., Ancrenaz K. & Andau P. 2005. Aerial surveys give new estimates for orangutans in Sabah, Malaysia. PLoS Biology 3(1): www.plosbiology.org
Arrhenius, O. 1921. Species and Area. Journal of Ecology. 9:95-99.
Baillie J.E.M., Hilton-Taylor C. & Stuart S.N. 2004. 2004 IUCN Red List of Threatened Species. A Global Species Assessment. IUCN, Gland, Switzerland and Cambridge, U.K.
Best, P.B. 1990. Trends in the inshore right whale population off South Africa, 1969-1987. Marine Mammal Science 6: 93-108.
Buchanan, G., J. Pearce-Higgins, M. Grant, D. Robertson, and T. Waterhouse. 2005. Characterization of moorland vegetation and the prediction of bird abundance using remote sensing. J. Biogeography, 32: 697-707.
Dinerstein, E., C. Loucks, A. Heydlauff, E. Wikramanayake, G. Bryja, J. Forrest, J. Ginsberg, S. Klenzendorf, P. Leimgruber, T. O’Brien, E. Sanderson, J. Seidensticker, and M. Songer 2006. Setting Priorities for the Conservation and Recovery of Wild Tigers: 2005-2015. A User’s Guide. WWF, WCS, Smithsonian and NFWF-STF, Washington, D.C, New York.
Henebry, G. M., & Merchant, J. W. 2002. Geospatial data in time: limits and prospects for predicting species occurrences. In Scott, J. M., Heglund, P. J., Morrison, M. L., Haufler, J. B., Raphael, M. G., Wall, W. A., & Samson, F. B. (Eds.), Predicting species occurrences: Issues of accuracy and scale. (pp. 291-302). Washington: Island Press.
Hochberg, E.J. and M.J. Atkinson. 2000. Spectral discrimination of coral reef benthic communities. Coral Reefs 19:164-171.
Gould.W. 2000 Remote Sensing of Vegetation, Plant Species Richness, and Regional Biodiversity Hotspots. Ecological Applications, 10, (6):1861-1870
Jiang Hong, J.R. Strittholt, P.A. Frost, and N.C. Slosser. 2004. The classification of late seral forests in the Pacific Northwest, USA using Landsat ETM+ imagery. Remote Sensing of Environment, 91:320-331.
Kerr, J.T. and M. Ostrovsky. 2003. From space to species: ecological applications for remote sensing. Trends in Ecology and Evolution, 18:299-305.
Key T., T.A. Warner, J.B. McGraw, and M.A. Fajvan. 2001. A Comparison of Multispectral and Multitemporal Information in High Spatial Resolution Imagery for Classification of Individual Tree Species in a Temperate Hardwood Forest. Remote Sensing of Environment 75: 100-113.
Koy, K., W. J. McShea, P. Leimgruber, B. N. Haack, and Myint Aung. 2005. Percent canopy cover using Landsat imagery to delineate habitat for Myanmar’s endangered Eld’s deer (
Cervus eldi). Animal Conservation 8:289-296.
Leimgruber, P., C.A. Christen, A. Laborderie. 2005. The impact of Landsat satellite monitoring on conservation biology. Environmental Monitoring and Assessment 106: 81-101
Liu, J., M. Linderman, Z. Ouyang, L. An, J. Yang, and H. Zhang. 2001. Ecological degradation in protected areas: the case of Wolong nature reserve for giant pandas. Science 292: 98-101.
Linderman, M., J. Liu, J. Qi, L. An, Z. Ouyang, J. Yang, and Y. Tan. 2004. Using artificial neural networks to map the spatial distribution of understory bamboo from remote sensing data. International Journal of Remote Sensing 25, 1685-1700.
Loh, J., R.E. Green, T. Ricketts, J. Lamoreux, M. Jenkins, V. Kapos, and J. Randers. 2005. Living Planet Index: using species population time series to track trends in biodiversity. Phil. Trans. R. Soc. B 360: 289-295.
Loucks, C.J., L. Zhi, E. Dinerstein, W. Dajun, F. Dali, and W. Hao. 2003. The giant pandas of the Qinling mountains, China: a case study in designing conservation landscapes for elevational migrants. Conservation Biology 17: 558-565.
Manley, P.N., W.J. Zielinski, M.D. Schlesinger, and S.R. Mori. 2004. Evaluation of multiple-species approach to monitoring species at the ecoregional scale. Ecological Applications 14: 296-310.
McGraw, J.B., T. A. Warner, T. L.Key, and W. L. Lamar. 1998. High spatial resolution remote sensing of forest trees. Trends in Ecology and Evolution 13: 300–301.
McShea, W. J., P. Leimgruber, Myint-Aung, S. L. Monfort., and C. Wemmer. 1999. Range collapse of a tropical cervid (
Cervus eldi) and the extent of remaining habitat in Central Myanmar. Animal Conservation 2:173-183.
Milner-Gulland, E.J., M.V. Kholodova, A. Bekenov, O. Bukreeva, I.A. Grachev, L. Amgalan, and A.A. Lushchekina. 2001. Dramatic declines in saiga antelope populations. Oryx 35: 340-345.
Mumby, P.J., E.P. Green, A.J. Edwards, and C.D. Clark. 1997. Coral reef habitat mapping: how much detail can remote sensing provide? Marine Biology 130:193-202.
Murn, C., M.D. Anderson, and A. Anthony. 2002. Aerial survey of African white-backed vulture colonies around Kimberley, Northern Cape and Free State provinces, South Africa. South African Journal of Wildlife Research 32:145-152.
Osborne, P.E., J.C. Alonso, and R.G. Bryant. 2001. Modeling landscape-scale habitat use using GIS and remote sensing: a case study with great bustards. J. Applied Ecology 38:458-471.
Raxworthy, Christopher J., Enrique Martinez-Meyer, Ned Horning, Ronald A. Nussbaum, Gregory E. Schneider, Miguel A. Ortega-Huerta & A. Townsend Peterson. 2003. Predicting distributions of known and unknown reptile species in Madagascar. Nature 426: 837-841.
Rushton, S.P., S.J. Ormerod, and G. Kerby. 2004. New paradigms for modeling species distributions? J. Applied Ecology, 41: 193-200.
Sanderson, E., J. Forrest, C. Loucks, J. Ginsberg, E. Dinerstein, J. Seidensticker, P. Leimgruber, M. Songer, A. Heydlauff, T. O’Brien, G. Bryja, S. Klenzendorf, and E. Wikramanayake 2006. Setting Priorities for the Conservation and Recovery of Wild Tigers: 2005-2015. The Technical Assessment. WCS, WWF, Smithsonian, NFWF-STF, New York – Washington, D.C.
Sauer, J. R., J. E. Hines, and J. Fallon. 2005. The North American Breeding Bird Survey, Results and Analysis 1966 – 2004. Version 2005.2. USGS Patuxent Wildlife Research Center, Laurel, MD.
http://www.mbr-pwrc.usgs.gov/bbs/bbs.html (accessed January 2007)
Sidle, J.G., D.H. Johnson, and B.R. Euliss. 2001. Estimated areal extent of colonies of black-tailed prairie dogs in the northern Great Plains. J. Mammology 82: 928-936.
Stauffer, D. F. 2002. Linking populations and habitats: Where have we been? Where are we going? In Scott, J. M., Heglund, P. J., Morrison, M. L., Haufler, J. B., Raphael, M. G., Wall, W. A., & Samson, F. B. (Eds.), Predicting species occurrences: Issues of accuracy and scale (pp. 53-61). Washington: Island Press.
Tracey, J.P., P.J.S. Fleming and G.J. Melville. 2005. Does variable probability of detection compromise the use of indices in aerial surveys of medium-sized mammals? Wildlife Research 32(3) 245–252.
Turner, W., S. Spector, N. Gardiner, M. Fladeland, E. Sterling, and M. Steininger. 2003. Remote sensing in biodiversity science and conservation. Trends in Ecology and Evolution 18(6): 306-314.
Whitehouse, A.M., A.J. Hall-Martin, and M.H. Knight. 2001. A comparison of methods used to count the elephant population of the Addo Elephant National Park, South Africa. African Journal of Ecology 39: 140-145.