Chapter 4. Trends in Selected Biomes, Habitats, and Ecosystems: Forests
Authors:
James Strittholt, Conservation Biology Institute
Marc Steininger, Conservation International
Contributors:
Colby Loucks, World Wildlife Fund (WWF-US)
Ben White, Global Land Cover Facility, University of Maryland
Reviewers:
Mette Løyche Wilkie, Food and Agriculture Organization of the United Nations
Manuel Guariguata, Center for International Forestry Research
Remote sensing based indicators for forests:
- Extent of component ecosystems
- Forest change
- Rate of deforestation/reforestation
- Forest intactness
- Area and number of large forest blocks
- Forest fragmentation
- Carbon storage
- Area and location of old-growth forests
- Area and location of plantations
- Forest degradation
- Area and location of sustainable forestry
- Alien species
- Fire occurrence
The physical characteristics of forest ecosystems reflect sunlight in the visible, near-infrared, and middle-infrared regions of the light spectrum in ways that are easily distinguished from other types of vegetation cover. Thus, remote sensing has become an important tool for evaluating forest ecosystems at multiple spatial and temporal scales to examine and monitor forest composition, structure, and function (Kerr and Ostrovsky 2003).
4.1 Delineating Cover and Estimating Change in Extent
Leaves are full of pigments that absorb visible light. Therefore, because of their dense leaf cover, forested areas tend to reflect less light from the visible region of the spectrum (and thus appear darker than) other vegetation types. In contrast, leaves strongly reflect near-infrared light — which is not visible to our eyes — and thus they appear brighter when near-infrared data are mapped. Water in forest leaves absorbs near-infrared light, and thus forests tend to appear dark when middle-infrared images are displayed. The middle-infrared is also best able to reveal “canopy shading.” This is caused by the canopy geometry (i.e., the uneven tree and branch height) of forests. In general, taller forests with more uneven canopies, such as old-growth forests, appear darker in the middle-infrared than shorter forests with more even canopies, such as young secondary regrowth, because of a combination of canopy shading and the water absorption of leaves. This darkening trend is also seen in the near-infrared, although less so. Inundated forests usually appear even darker still, because of even more light absorbed by standing water or wet soils beneath the canopy. The brightness in all of these spectral regions also depends on the colour of the background underneath the canopy, which is a mix of soil and vegetative litter. The influence of the background colour is more obvious when leaf cover is lower, such as in areas of more open vegetation during the dry seasons when leaves have senesced.
When observing optical images of forests and other types of vegetation cover, it is useful to consider how green they are (leaf cover), how bright they are (canopy shading), and how wet they are (inundation). As an example, we can consider the changes in reflectance in these three spectral regions as a forest regenerates, developing from mostly shrub cover with relatively little leaf cover and an even canopy to mostly young trees with more leaf cover and a somewhat even canopy and then to a tall forest with maximum leaf cover and a very uneven canopy (Moran et al. 1994; Steininger 1996). As a forest grows, the visible reflectance declines rapidly to as low as 5 percent when leaf-area indices are 4 or greater. Near-infrared reflectance increases in the early stages of regrowth because of increasing leaf cover, but then often decreases somewhat at later stages of regrowth because of canopy shading. Throughout the regrowth sequence, middle-infrared tends to gradually decline. Depending on the type of forest and the rate of regrowth, older secondary forest regrowth begins to appear similar to old-growth forest.
Another characteristic of optical images, at least those at finer spatial resolutions, is the texture of the image. Images of some forest types appear speckled or mottled. For example, a tall tropical forest appears speckled in Landsat images because the spatial resolution, 30 metres, is close to the size of a single tree crown. Thus one pixel may image a tree crown (bright in near-infrared), and its neighboring pixel would image a shaded gap in between two crowns (dark in near-infrared). In a coarser image, such as a MODIS reflectance image, this same forest would appear smooth. A mottled texture in a Landsat image, in which the image appears to have small clumps of several pixels that are brighter than neighboring clumps, may indicate a logged forest that has gaps the size of several tree widths.
Global maps from 0.5-kilometre to 1-kilometre resolution satellite data already provide a reasonable snapshot of the general extent of forest cover. The
Global Land Cover 2000 (Bartholomé and Belward 2005) map product was based on 1-kilometre spatial resolution satellite data acquired over the whole globe by the VEGETATION instrument on board the SPOT 4 satellite). The GLC2000 product includes eight forest classes: (1) broadleaved, evergreen; (2) broadleaved, deciduous, closed; (3) broadleaved, deciduous, open; (4) needle-leaved, evergreen; (5) needle-leaved, deciduous; (6) mixed leaf type; (7) regularly flooded, fresh water; and (8) regularly flooded, saline water (Bartholomé and Belward 2005). The reported overall accuracy of this product is greater than 70 percent (Mayaux et al. 2006). A similar product, the
Global Land Cover Characteristics Database (GLCCD), was created from a 1-kilometre spatial resolution Advanced Very High Resolution Radiometer (AVHRR) and likewise contains coarse-level forest classes (Loveland et al. 2000).
The coarse spatial resolution of these global products provides a useful, general baseline of forest cover by general forest type, but they are not suitable for global or national forest operational monitoring because of insufficient accuracy and the lack of detailed forest themes. For example, small nonforest patches within forest dominated regions and small forest patches in low forest regions remain undetected with coarse sensors, leading to over- and underestimation of forest extent. Some of this error can be reduced by calibration against high-resolution data (Mayaux and Lambin 1997), but with the widespread availability of moderate- and high-resolution sensors, it is better to conduct national and subnational forest mapping using these higher resolution platforms. For not only is spatial accuracy of forest extent enhanced, but also these sensors allow for better discrimination of forest types and other characteristics such as forest age.
Spatial resolution is enhanced at the global level with more refined sensors like
MODIS (250- to 500-metre spatial resolution) and
MERIS (300-metre spatial resolution), which were developed to facilitate global change assessments. The MODIS land cover product used a supervised classification approach in which training sites were provided using a decision-tree classifier (Friedl et al. 2002; McIver and Friedl 2002). Map accuracy for version 3 of this global-mapping product has been reported to be 75–80 percent overall; 70–85 percent by continental regions; and 60–90 percent for individual cover classes (Friedl 2007). MODIS Vegetation Continuous Fields (or Percent Tree Cover) has provided additional land cover detail specific to forests (Hansen et al. 2003), advancing our ability to map forests at the global level with greater precision and accuracy.
These coarse spatial resolution products have considerable value at the global scale, but they lack sufficient thematic and spatial detail needed for habitat assessments on the level of individual countries, and the spatial resolution is too coarse for a basis for high-quality forest-change monitoring. Nevertheless, technical remote sensing capabilities have advanced dramatically over the past decade, and monitoring forests at the national level is now a feasible goal for most countries (Mollicone et al. 2003; DeFries et al. 2006).
Moderate resolution sensors, including
Landsat,
ASTER,
SPOT HRV, and
IRS with spatial resolutions from 15 to 60 metres have formed the foundation for forest mapping at the national and subnational level. Numerous examples of results (many available online) from these efforts are provided in the resource section at the end of this chapter. For most nations, employing these sensors to conduct their forest monitoring needs provides the most cost-effective method, with reasonable thematic and spatial accuracies (usually greater than 80 percent) routinely observed. Also, there is a growing digital image library for many of these sensors, allowing for informative change detection assessments and trends analyses.
There are newer, high-resolution optical sensors (5-metre resolution or less) now being used to map and evaluate forest ecosystems. A number of airborne sensors collect digital imagery at multiple spectral bands such as CASI (Anger et al. 1994), as well as satellite platforms such as
IKONOS and
QuickBird. These sensors often provide enough spatial and spectral detail to map individual trees. High-resolution remote sensing plays a minor or nonexistent role when monitoring forests at the global or national scale: high-resolution imagery is generally cost-prohibitive from the standpoints of both data acquisition and handling. However, particular forest community types or individual tree species of global importance that are distinctive (e.g.,
Dipterocarpis spp. in Thailand and
Doona congestiflora in Sri Lanka) could be monitored using these technologies.
Active remote sensing systems (radar and — to a lesser extent — lidar) are emerging as the new generation of forest remote sensing tools because they provide a number of advantages over optical systems: (1) ability to collect data in regions of continuous cloud cover; (2) usefulness in tracking forests that are seasonally inundated by water; (3) better estimates of forest biomass because they provide a 3-D perspective of the forest; (4) help in the classification of stand age; and (5) measurement of subcanopy characteristics (Wulder et al. 2004). Like all remote sensing technologies, active sensors have technical disadvantages as well (e.g., radar does not perform particularly well in hilly terrain), and much more research is needed before we realize the full benefits. Active remote sensing is still cost-prohibitive for large areas such as many national assessments, but may have a place in a global-/national-monitoring context in the near future or for special cases today. Finally, researchers are exploring data integration for mapping forests. For example, by combining Landsat Thematic Mapper imagery with lidar (Lefsky et al. 2002) or radar (Treuhaft et al. 2004), a third dimension to otherwise two-dimensional forest mapping can be achieved.
4.1.1 Mapping humid tropical forests Under ideal conditions, mapping forests using remote sensing is reasonably accurate and cost-effective. However, different forest biomes pose unique challenges to remote sensing. The most obvious technical issue for humid tropical forests is the difficulty in obtaining clear images of regions that are persistently cloudy, such as most rainforest and almost all montane, submontane, and island forest regions. In mountainous areas, rugged topography adds further complexity to image interpretation, primarily because of shading, which in some areas can be quite severe. Because optical sensors rely on reflected light, shaded areas reflect little or no light back to the sensor, resulting in little or no reflectance data used to discriminate different cover classes. Under variable illumination conditions, the same forest type appears differently in satellite images, depending on whether the forest is on the sunlit or shaded side of a mountain. Under heavy shading conditions, as much as 10–25 percent of a region can be lost or compromised by shadows.
4.1.2 Mapping dry tropical forests Because of their heterogeneous pattern, loss of leaves in the dry season, and variability of the tree canopy, tropical dry forests are susceptible to misclassification without validation with ground information. Great care should be taken in choosing when satellite images are acquired: — both wet versus dry seasons as well as the particular conditions during any given year. Ideally, for each time period of interest, two sets of images — one from the end of the wet season and one from the end of the dry season — should be processed. This will allow for the mapping of forest extent (end of wet season imagery) and the degree to which the forests retain their leaves (end of dry season). From this information, it is possible to assess the percentage of canopy cover and to better separate the region into discrete classes based on spectral reflectance.
Also, many dry forests frequently burn as the dry season progresses and may begin to resemble bare lands or dry agricultural fields during this period. This underscores the need for a wet season image, which will clearly demonstrate the extent of forests, agricultural areas, and residential areas.
Degraded areas, often found in the wetter, more semievergreen dry forests, may resemble mixed deciduous forests in satellite data. Also, care should be taken to properly assess areas that have been degraded against those areas that have a natural open canopy system. One clue is that degraded areas are more likely to be found closer to human settlements or roads. Dry season imagery may contain open canopies, but also an understory filled with vegetation such as bamboo. Riparian forests will likely be similar to evergreen or mixed deciduous forests, especially in a dry-season image. It is likely that they contain different tree species, despite the similarity in phenology.
4.1.3 Mapping boreal and temperate forests The overall principles for interpreting optical images of forests for brightness, greenness, and wetness apply to both temperate and boreal forests. For boreal forests, it can be particularly useful to estimate varying levels of inundation because many of the boreal forests are closely associated with extensive wetlands. Boreal forests are also characterized by cycles of forest regeneration resulting from recovery following wildfires and logging. Thus, estimating the age, height, biomass, or stage of regeneration can be important. The rate of forest growth is slower in the boreal zone than in other zones, so it takes longer for secondary forests to appear similar to mature forests. This is further emphasized by the typical succession from shrubs to broad-leaved trees to needle-leaved trees. The different shapes and distributions of leaves in canopies mean that broad-leaved forests tend to appear much brighter and greener than needle-leaved forests. In the boreal context, areas that appear brightest and greenest can be either mature hardwoods or early regenerating conifer stands, which are often mixed with deciduous species.
Remote sensing using finer-resolution sensors (e.g., Landsat and ASTER) is the most effective and economical approach to mapping boreal forest communities (e.g., black spruce, jack pine, and larch) that would be most meaningful for biodiversity purposes, but because of a relative lack of commercial interest in the more remote regions of the boreal, it has not been a high mapping priority for Governments. However, this is changing as human development rapidly encroaches into the undeveloped regions of the boreal zone. Community-level data is important for two reasons: (1) some boreal forest types are rarer or more threatened than others (e.g., riparian white spruce forests and white pine forests), and these communities should be monitored more carefully; and (2) the goal of representing biodiversity in a system of global protected areas can be achieved only when we know the extents and conditions of the different forest communities. Mapping the extent of temperate deciduous forests can be accomplished with high levels of accuracy when there is high contrast between the forest and other cover types. It becomes more challenging when forest and shrublands are intermixed. In drier regions, deciduous forest may gradually transition to open woodlands, and it is difficult to consistently define a border between the two. Furthermore, the season of the year and different precipitation levels can make appearance of both forest and shrublands vary dramatically. The best way to distinguish varying levels of deciduousness, to characterize gradients of forest and woodland types, is to analyse images from different seasons.
4.1.4 Change in forest extent An important source for forest change information is the
United Nations Food and Agriculture Organization (FAO) series of
Forest Resources Assessments (FRAs) produced from national data.
FRA 2000 and
FRA 2005 are the two most recently published reports, with another one planned for 2010. FRAs have traditionally been produced using field data; however, FRA 2000 included an assessment of forest area changes in the tropics based on remote sensing. Increased use of remote sensing technologies is being promoted for the next Forest Resource Assessment report scheduled for 2010 (FAO 2006). It is anticipated that the FRA for 2010 will include an analysis of a sample of Landsat data (30-metre resolution data): a 10-kilometre by-10-kilometre sample of multi-temporal images (1975-1990-2000-2005) located at every 1-degree-by-1-degree latitude/longitude intersection (FAO 2006). These new estimates should provide more accurate and consistent results and should represent one of a suite of global estimates of trends using varying sources of satellite data.
Despite very different data sources and methods, estimates of the global rate of deforestation during the 1990s derived from remote sensing, and FAO results derived largely by other means agree to within 15 percent. This is encouraging for global research. However, the level of agreement declines when these products are compared at the continental level or for shorter time periods. For example, the same estimates show a 40 percent disagreement at the continental level (Global Observation of Forest and Land Cover Dynamics 2004). The estimates of deforestation rates for individual countries reported to FAO can differ by as much as 100 percent from estimates derived from wall-to-wall mapping with moderate-resolution data (Tucker and Townshend 2000; Steininger et al. 2001). Much of this disagreement is attributable to the following three factors: 1) inability to detect small area changes with medium to coarse resolution remote sensing, 2) inadequate resources and capacity to collect timely and comparable national-level information over time – especially in Africa, and 3) differences in the definition of “forest”– partly related to the choice of methodology both between countries and over time within countries. The latter problem reinforces the importance of clear standards and the clear understanding of definitions and data-handling methods.
Other examples of global forest change efforts include (1) a coarse resolution (8-kilometre resolution product) created by the University of Maryland (Hansen and Defries 2004); (2) a global sample of moderate-resolution (30-metre) images, analysed by the European Commission’s Joint Research Centre (Achard et al. 2004); and (3) numerous country- or region-level change analyses carried out by researchers from a variety of academic, governmental, and nongovernmental organizations. Two examples of NGO efforts are highlighted as case studies below.
Case Study 4.1: Forest Cover Change in Paraguay
Author: Ben White, Global Land Cover Facility (GLCF), University of Maryland
Indicator: tropical forest extent and change
Potential monitoring scales: landscape, small and large nations
Sensor: Landsat TM and Landsat Enhanced Thematic Mapper (ETM+)
Imagery cost/hectare: free
a. Introduction The moist tropical forests, tropical grasslands, and savannas that make up the Atlantic Forest are one of the world’s most diverse and endangered ecosystems. The forest has diminished to roughly 10 percent of its pre-Columbian extent, largely because of pressures from farming, logging, and population growth. Nevertheless, the remaining habitat is astonishingly diverse, with more than 2,000 estimated vertebrate species and more than 20,000 plant species. By the early 1970s, the Brazilian interior Atlantic Forest was almost completely destroyed, while portions of the Atlantic Forest remained intact in Argentina and Paraguay. Although Brazil and Argentina established parks around Iguazú Falls to preserve portions of the forest, the majority of the Atlantic Forest in Paraguay has been almost entirely eliminated or fragmented. Today, the remaining Paraguayan Atlantic Forest is largely a function of accessibility and protected status. Although circumstances leading to the post-1970 deforestation are complex, three significant regional dynamics are recognized. The construction of a dam at the northern tip of the Paraná River substantially impacted the watershed, which was tied to traditional land dynamics throughout most of Paraguay. The Central Bank of Paraguay and the Inter-American Development Bank (IADB) have determined that in recent years, there has also been an increasing lack of governance throughout the border departments (states), making environmental management increasingly challenging. Perhaps the most significant driver of land cover change was the introduction of mechanized soy agriculture. In Alta Paraná and Itapua departments, the correlation of deforestation with soy production was particularly strong.
b. Methods The methodology for the Paraguayan forest cover change mapping was initiated with the coregistration of orthorectified Landsat TM with ETM+ Level1G imagery. A data mask was then developed, incorporating only those pixels that need to be classified. The masking was followed by a standard ISODATA clustering of the multitemporal Landsat imagery. A binary training mask was created for each class. The spectral clusters resulting from the ISODATA clustering were then labeled using an automated application developed by GLCF. The masking/labelling routine was repeated until all pixels were classified. As needed, the GLCF personnel performed any required editing and filtering. The classified tiles were then mosaicked into a single country product.
c. Results The GLCF became involved with the Paraguayan conservation organization, Guyra Paraguay, in an effort to map the changing land cover and to help guide local, national, and NGO managers and stakeholders. GLCF staff was able to map the vegetation changes by using Landsat satellite imagery and the methods outlined above, while Guyra Paraguay provided the ground-truthing necessary to validate the GLCF results (figure 4.1).

Since then, Guyra Paraguay has conducted its own updating of these deforestation maps, using MODIS data, and published the results in a national newspaper. This was used in a geographic information system (GIS) to create edge-area maps of each forest patch. The change over time in this ratio is reported per department. All of these data are available online at
GLCF
Case Study 4.2: Dry Tropical Forest Mosaic of Eastern Cambodia
Author: Colby Loucks, World Wildlife Fund US (WWF)
Indicator: dry tropical forest extent
Potential monitoring scales: landscape, small and large nations
Sensor: Landsat TM and Landsat ETM+
Imagery cost/hectare: free
Limitations on accuracy: seasonality, fires, ground-truth data
a. Introduction The dry forests of eastern Cambodia are the largest contiguous area of dry forest in all of mainland Southeast Asia. Cambodia’s eastern plains are dominated by deciduous and dry dipterocarp forests, interspersed with patches of semievergreen forest. Many large mammal species require all three of these forest types to hunt or breed sometime during the year. A study by the WWF produced an estimate of the distribution and change in extent from 1990 to 2002 of three types of forest: semievergreen, mixed deciduous, and dry dipterocarp. It estimated how much of each type was converted to agricultural, residential, or barren land cover.
b. Methods The analysis was based on two Landsat images of the same area, both from the dry season: December 21, 1990, and February 13, 2002. WWF collected ground-reference data during surveys of the southeast portion of the study area. WWF also collected GIS data on topography, roads, villages, protected areas, and rivers to provide contextual information. These data were supplemented with interviews with local experts.
The analysis of the two images began with geometric registration of the 2002 image and then coregistration of the 1990 image to the 2002 image. This was achieved with subpixel accuracy (less than 30 metres). The two images, each with six data layers, one for each reflectance channel, were then combined to create a single two-date file with 12 data layers. This file was then used in an unsupervised classification to produce 250 classes, or clusters of data with greatest similarity among the 12 layers. These classes were then labeled (i.e., assigned to one of several possible cover types), referring to the suite of reference and supplemental data collected. Additional field surveys were conducted in areas where interpretation of the raw images was most difficult and where classification results were most suspect. The latter was evidenced by areas where several classes were interspersed in the results and where results appeared to conflict with experts notes. The data from the second survey were used to refine the initial classification. The final map was then filtered with a 3x3 grid cell box neighborhood focal filter. This filter returns the majority value to the centre pixel and filters out isolated pixels.
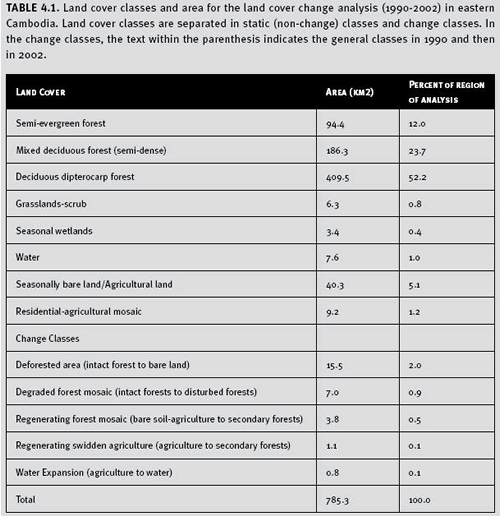
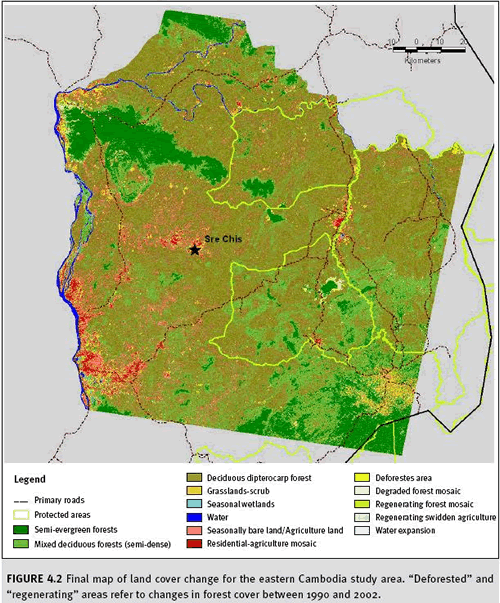
c. Results Mapped results of the forest change analysis for Cambodia are presented in Figure 4.2. We found that 2.9% of the dry dipterocarp, mixed deciduous, and semi-evergreen forests in the study area were deforested over the 12 year time period (Table 4.1). The majority of the forest loss occurred near villages that existed in 1990. Expansion of these populated areas occurred over the 12 year period with the conversion of most of the forest to residential-agricultural lands. Additional forest loss was also found along road networks, several of which were created or improved since 1990. The semi-evergreen forests were deforested as a result of both conversion to swidden agriculture and deforestation along road networks that were built through these blocks of forests. We also found that 0.6% of the forests in the study area had regenerated (Table 4.1).
The majority of the regeneration areas were from prior intensive timber operations. A small portion of the regenerating forest was from abandoned swidden agricultural lands found in semi-evergreen forests. Therefore we measured a net decrease in forest areas of 2.3% in the study area over the 12 year time frame. A majority of the study area still contains native land cover (89.1%), while approximately 9%of the study area has been converted to human-dominated land uses (i.e. residential or agricultural lands).
Table 4.1. Land cover classes and area for the land cover change analysis (1990-2002) in eastern Cambodia. Land cover classes are separated in static (non-change) classes and change classes. In the change classes, the text within the parenthesis indicates the general classes in 1990 and then in 2002.
 d. Limitations
d. Limitations As with many remote sensing analyses, knowledge of the region and forest types greatly increases the accuracy of the classification. This is especially the case with the dry deciduous forest mosaic found in eastern Cambodia. During the dry season, dry dipterocarp forests may resemble agricultural or seasonally bare areas in Landsat imagery. Burned agricultural lands may resemble naturally burned dry dipterocarp forests. Riparian forests resemble mixed deciduous or semievergreen forests, but are likely made up of different tree species.
Information concerning the location of permanent water sources, elevation, and fire activity is also important. Areas with perennial water supply are more likely to have semievergreen or mixed deciduous forests. The dry forest mosaic of eastern Cambodia is frequently burned, both by wildfires and by humans to stimulate growth of new plants. Burning frequency and extent confound land cover classification, with the potential to misclassify dry dipterocarp forests as “seasonally bare land.” Recently burned land, whether it is an agricultural or natural area, has similar spectral signatures in the dry season. Firsthand knowledge of all these areas is likely to improve the final classification. The seasonality of this forest type argues for obtaining imagery for both the wet and dry seasons. Doing so will assist in classification by allowing comparison of seasonal signatures for each time period and determination of seasonality versus time-period changes in land cover. This also helps minimize misclassification among nonforest and dry forests. Our initial supervised classification misrepresented the mixed deciduous forest class in the region. However, ground-reference data revealed an understory of bamboo. Thus, field verification was important for image interpretation and to accurately classify the mixed deciduous forest class. We also identified a number of areas in the semievergreen forests that were cleared for roads or small clearings that were misclassified as “mixed deciduous” or “dry dipterocarp” forests.
Because of the seasonal differences and proclivity of fires in dry tropical forests, spectral signatures differ considerably for the same habitat type. A constant challenge is to discern changes that are permanent, seasonal, or temporary. Most projects do not have funds for extensive field verification, especially in remote and inaccessible regions such as eastern Cambodia. However, local knowledge becomes of paramount importance for an accurate land cover classification.
4.2 Forest Quality
A number of forest indicators fall under the heading of forest quality, including (1) forest intactness, (2) large forest blocks, (3) forest fragmentation, (4) area and location of old growth, (5) area and location of plantations, (6) changes in forest pests and diseases, and (7) fire occurrence. Remote sensing has a role to play in assessing all of these indicators, and each will be briefly addressed below.
4.2.1 Forest intactness and large forest blocks We know that natural forest landscapes lose components and functionality as human uses expand and continue over time. There exists a continuum of forest quality or “intactness,” ranging from a totally pristine environment to a totally developed environment bereft of native species. Quantifiable and replicable indices and scales of measurement are needed to score forest landscapes on this continuum, and this is an active area for many conservation NGOs because of the importance of these forested areas to biodiversity. While methods have yet to be fully standardized, mapping of forest intactness continues to be carried out by a number of conservation organizations throughout the world. Monitoring forest intactness remains a conservation imperative and a valuable indicator for global and national forest monitoring.
An intact forest landscape does not necessarily consist of old trees and may not even be entirely forested. Simply stated, intact forest landscapes are contiguous mosaics of natural habitat types (forest and nonforest alike) in forest-dominated regions that either have never been subjected to industrial human activities or have sufficiently recovered from such activities in the past to the point where the composition, structure, and function of the forest landscape are relatively complete (Strittholt et al. 2006). In some localities (e.g., portions of Amazonia, Canada, Central Africa, Indonesia, Russia, and the United States), these are the remaining “frontier forests” as defined and mapped at a very coarse level by Bryant et al. (1997). In other localities, these are forest landscapes that have recovered from previous human disturbance to the point where they possess many, if not most, of their original forest characteristics (e.g., portions of the eastern United States and Canada, Eastern Europe, and portions of Asia). Also, it is important to note that intact forests are not static systems. On the contrary, an ecosystem with a high level of intactness is one that is able to maintain its biodiversity and ecosystem functionality over time—not in any fixed, quantitative sense, but rather as a dynamic property (O’Neill et al. 1986; Holling 1992). To a large extent, many of the compositional (e.g., types of species present) and structural (e.g., size of trees and complexity of forest canopy) components of forest landscapes are size-independent. However, home range needs for some animal species and many ecological processes (e.g., natural regeneration, natural disturbance, nutrient cycling, predator-prey interactions, and migration and dispersal) require considerable areal extents to operate within their natural range of variability. Therefore, the definition of intact forest landscapes requires a reference to some minimum size. Natural disturbance regimes (e.g., fire, windthrow, and phase gap dynamics) and areal requirements of native, large home range species have been used repeatedly to help establish ecologically meaningful size thresholds for intactness. To date, the
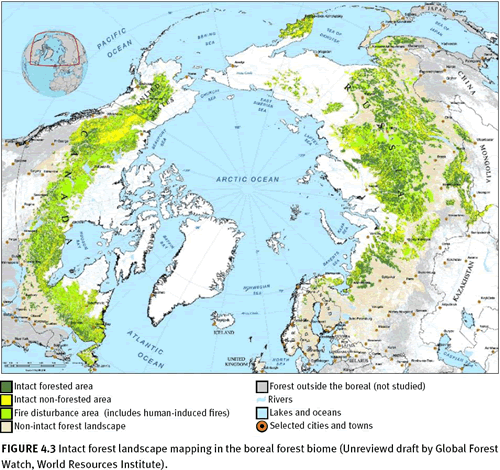
Global Forest Watch (GFW) network, within the
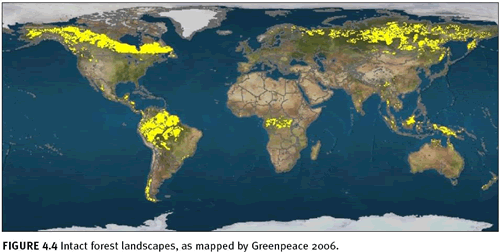
World Resources Institute, has advanced the mapping of intact forest landscapes more than any other organization. Using moderate resolution satellite remote sensing, GFW partners have mapped and continue to update forest intactness products for Brazil (Barreto et al. 2006), Canada (Smith et al. 2000; Lee et al. 2003; Lee et al. 2006), Central Africa (Minnemeyer 2002; Van de Pol et al. 2005), Chile (Neira et al. 2002), Indonesia (Achmaliadi et al. 2002), Russia (Yaroshenko 2001; Aksenov et al. 2002; Aksenov et al. 2006), the United States (Noguerón 2002; Strittholt et al. 2006), and Venezuela (Bevilacqua et al. 2002). In 2002, GFW partners working in the boreal biome compiled existing forest intactness maps to form a panboreal map for the world (figure 4.3), and Greenpeace created a global forest intactness map based on standardized rules for mapping forest intactness thresholds (figure 4.4; Greenpeace 2006). Based on a number of different remote sensing data sources, this study concluded that about 13 million square kilometres remain as intact forest landscapes (23 percent of the forest zone and 9 percent of the earth’s surface). The remainder of the forest zone was identified as degraded, converted to plantations, or fragmented to areas smaller than 500 square kilometres in size by roads, settlements, etc. This global overview somewhat underestimates intactness because of the scale and definition of constraints, but it does provide a valuable global overview. The majority of the remaining intact forest landscapes of the world are located in the boreal/taiga forests of Alaska, Canada, and Russia (43.8 percent) and dense lowland tropical forests of the Amazon, Congo, and Southeast Asia Pacific (49 percent). The remaining intact forests are scattered widely among the other forest biomes. Of the intact forest landscapes, only 8 percent lie in strictly protected areas (IUCN categories I–III), according to the
United Nations Environment Programme (UNEP)/
IUCN World Database on Protected Areas (CBI unpublished report 2006). Of all countries full or partly within the forest zone, 82 (over half) have lost all of their intact forest landscapes. Of the remaining countries, half again have less than 10 percent of their forests still intact. Only 14 countries control more than 90 percent of the world’s remaining intact forest landscapes, led by Canada, Brazil, Russia, Papua New Guinea, the Democratic Republic of the Congo, and Indonesia. The monitoring of large forest blocks by country or by ecoregion is a valuable forest condition indicator that can be effectively achieved using moderate resolution remote sensing. These large blocks are important ecologically because they are the areas most likely to harbor native biodiversity and natural ecological and evolutionary function. With a monitoring process in place, these blocks can be tracked for change, and where they are being dissolved by human encroachment and use, we can anticipate negative ecological consequences. Monitoring forest intactness and large forest landscape blocks is a special subset of a more traditional national forest survey. Assessing intactness could be tied directly to national forest surveys or be handled separately.
 4.2.2 Forest fragmentation
4.2.2 Forest fragmentation The destruction and fragmentation of natural habitats (including forests) is widely reported as the most significant driver in the global decline in biodiversity. Riitters et al. (2004) created digital forest fragmentation maps from GLCC land cover maps (AVHRR, circa 1992) where each pixel value represents a forest fragmentation category for the surrounding 81 square kilometre landscape. Repetition of this mapping assessment would provide a general global overview and highlight coarse spatial trends, but it will be unsuitable for addressing most biodiversity concerns resulting from forest fragmentation: ecological impacts on species operate at finer scales. Mapping forest fragmentation has been demonstrated using moderate-resolution imagery and ancillary data for entire countries such as the United States (Heilman et al. 2002; Riitters et al. 2002), which could be applied to other nations regardless of their size. (More in-depth discussion, including issues of data, metrics, standards, and scale in mapping fragmentation can be found in
Chapter 10)
4.2.3 Area and location of old-growth forests Because of the importance of old-growth forests to native biodiversity and its continuing decline, it is important in any global/national survey strategy to monitor where these forests exist and how they change over time. While still an area of active research for remote sensing, currently age (or area of old growth) in some forest community types can be mapped with moderate to high spatial and thematic accuracy. Most studies on mapping forest age (including old growth) have been conducted in temperate forest ecosystems, especially those dominated by conifers. For example, temperate rainforests have been successfully mapped based on moderate-resolution remote sensing data using a number of different analytical techniques. Cohen et al. (1995) used unsupervised classification to map discrete forest age classes in the U.S. Pacific Northwest and later mapped the same region modelling continuous age (Cohen et al. 2001). Ohmann and Gregory (2002) employed direct gradient analysis and nearest neighbor imputation to predict detailed site conditions. Jiang et al. (2004) mapped forest age in the same region, using a technique called “optimal iterative unsupervised classification,” which resulted in accuracies between 80–90 percent for broadly defined age classes (figure 4.5). Old-growth forests also have been mapped for portions of the boreal forests of Russia (Aksenov et al. 1999), montane regions of tropical America (Costa Rica: Helmer et al. Honduras: Aguilar and oak-pine forests of western Mexico (Lammertink et al. 2007). In almost every case, ancillary data (e.g., slope, elevation, roads, and ownership) were employed to help interpret remote sensing signatures. In tropical forests, multitemporal remote sensing data has been shown to be necessary (Kimes et al. 1998). Using moderate resolution imagery is the most cost-effective method for mapping forest age over relatively large areas, but the next-generation sensors are pushing the level of spatial and thematic detail even more.
There is a growing body of knowledge about forest age, using high-resolution sensors such as IKONOS and airborne platforms (Franklin et al. 2001; Clark et al. 2003; Nelson et al. 2003). Although the various techniques mentioned here have proven to be feasible and reliable for mapping forest age, the future of mapping this important forest characteristic will be based on either active sensors (e.g., radar or lidar) alone (Drake et al. 2002) or through a combination of these sensors with moderate- (e.g., Landsat TM) or high-resolution (e.g., IKONOS) imagery (Lefsky et al. 2002).
There are always pros and cons to every forest survey decision. For some forest biomes, it is feasible in terms of quality of the results and cost today to map forest age with reasonable accuracy with moderate-resolution imagery at the national or subnational extent. In situations where fine resolution and very narrow age classes are required, more effort, additional data, and more cost are also required. From the global perspective, narrowing the scope of work to regions of particular interest or biological importance over the near term is encouraged until technological advances and cost reductions are realized, allowing for more geographically broad application.
4.2.4 Area and location of plantations

Plantations continue to expand throughout the world, increasing by 2.8 million hectares per year during 2000–2005 (figure 4.6).
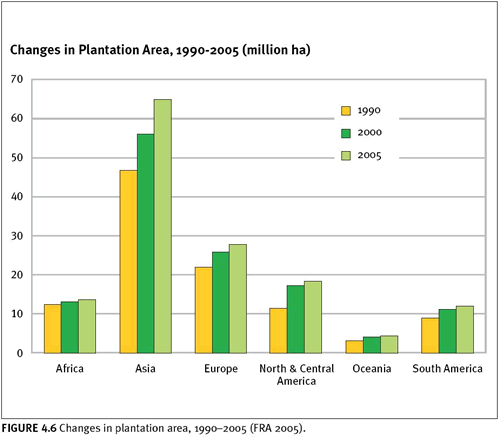
According to the definitions used by the most recent global forest assessment by FAO (FRA 2005), plantations are defined as a subset of planted forests consisting primarily of exotic species. Plantation forests are further subdivided into two classes: forests planted for wood and fiber production (or productive plantations) and forests planted for protected soil and water (or protective plantations). Of the approximately 140 million hectares of plantations in the world (3.8 percent of all forest cover), 78 percent have been identified as productive plantations and 22 percent as protective plantations (FRA 2005). Plantations are generally of lesser biodiversity value than natural forests, especially when comprised of exotic species (Hunter 1999). Their expansion warrants monitoring, especially when they replace high-quality native forests.
Because of their commercial value, plantations have been the focus of many remote sensing studies, usually over small geographic extents and for purposes other than biodiversity. Remote sensing studies of plantations have focused predominantly on mapping plantation characteristics such as timber volume (Trotter et al. 1997), age (Jensen et al. 1999; Ratnayake 2006), and productivity (Coops et al. 1998). Fewer studies have addressed distinguishing plantations from native forests directly; nevertheless, some insights can be gleaned from these studies. Evans et al. (2002) mapped land cover change, with particular interest in mapping the conversion of native forests to plantations for a portion of the southeastern United States, using a combination of Landsat TM and aerial photographs. Donahey (2006) used multispectral images to map forest plantations in the Atlantic Forest region of Costa Rica. SPOT 4 imagery and Landsat TM were used to map vegetation changes in Central Sumatra with reasonable success in mapping plantation cover types (Trichon et al. 1999). All of these studies confirm the importance of integrating field data and usually a mixing of data from different sensors, including high resolution imagery.
The use of moderate resolution images (20–30 metres) alone usually proves inadequate, unless the plantations cover very large areas. The UNEP World Conservation Monitoring Centre (2007) has divided forest plantations into four classes (temperate/boreal exotic species plantation, temperate/boreal native species plantation, tropical exotic plantation, and tropical native plantation) and mapped them at a coarse scale, using AVHRR-based satellite images. This data set is too coarse to help address most biodiversity monitoring questions. Data at this scale will only identify major changes in forest cover, such as forest clearance.
In regions where plantation species are dramatically different from the native forest cover (e.g., conifers planted where deciduous forests are the natural forest), the detection and monitoring of plantation extent and spatial configuration are relatively straightforward, employing moderate- and high-resolution remote sensing. However, in regions where the planted tree species are similar to the native forest, the differentiation of plantations from natural forests is far more difficult (and nearly impossible in some cases) without the aid of ancillary data. Furthermore, all of the other regional constraints that affect forest mapping (e.g., persistent cloud cover) apply here as well.
4.2.5 Fire occurrence Fire is the most common natural disturbance agent in forested ecoregions. Many forest species have evolved with fire, and some even require it for regeneration. Fires have a direct impact on local and regional biodiversity, which can be severely degraded under unnatural fire regimes. Poor forest management, human-caused ignitions, and (most important) climate change are fundamentally changing fire frequency, extent, and severity in many parts of the world. For that reason, monitoring forest fires is an extremely important indicator, and remote sensing provides an effective means for doing so (Fraser et al. 2000).
Satellite detection of fires now occurs in near-real time throughout much of the world (Tansey et al. 2004). Fires can be readily observed from optical satellite sensors.
AVHRR (Fraser et al. 2000) and
SPOT-VEGETATION (Grégoire et al. 2003) have been used to map burnt areas. Ash from fires is dark in all three optical regions, and burn scars can be observed if the burn was severe and recent enough and if the canopy is open enough.
MODIS, deployed in 2000, currently dominates fire monitoring at both global and regional scales (Justice et al. 2002). Fire mapping derived from MODIS is
available online with near-real-time mapping. These products show a very large number of fire ignitions in equatorial Africa and South America, with the largest fires in the boreal zone. Monitoring at these scales provides extremely useful information regarding ecological processes such as carbon storage and nutrient budgets, which is important to biodiversity, but this scale of monitoring does not provide enough detail on the intensity and detailed spatial configuration of fires, which is also very important from the standpoint of biodiversity. For example, MODIS imagery can delineate a general fire perimeter, but closer examination with a higher-resolution sensor (e.g., Landsat TM) reveals the configuration and extent of various severities (figure 4.7).

In this example, one can see areas of high severity (magenta) to areas untouched by the fire (green). There have been numerous studies integrating numerous sensors to understand the full impact of fires on natural systems (e.g., Steyaert et al. 1997).
Tracking fire and its impacts is one aspect of forest-fire monitoring. Another is to assess changes in levels of fire susceptibility. From a societal perspective, this provides an early-warning system to mitigate property damage and loss of human life. From a biodiversity perspective, understanding where forest landscapes are becoming more susceptible to fire, especially if it is outside the range of natural variability for a given region, provides spatially explicit guidance as to where regional biodiversity will be at risk.
4.3 Threats
4.3.1 Deforestation Systematic observations of the world’s forests have been ongoing since the 1990s. From the standpoint of monitoring deforestation in the climate change equation, remote sensing is the only practical approach (DeFries et al. 2006). Since the 1990s, changes in forest extent based on coarse-and moderate resolution satellite imagery have been monitored reliably for climate change modelling. Coarse resolution sensors can detect only large-scale clearings, while clearings of 0.5 hectare can be detected in Landsat images with 30-metre resolution.
Deforestation is not randomly distributed across the world (Tucker and Townsend 2000), so it is important to monitor some areas more intensely and at higher resolution, especially where there is high risk of occurrence. Sampling has been shown to be effective at estimating deforestation (Strahler et al. 2006). Therefore, the nonrandom nature of this human impact must be taken into account to develop an effective sampling strategy.
Figure 4.8 shows an example from Alberta, Canada where the impacts from clearcut forestry and oil and gas exploration have been dramatic over a 10-year period. More information on deforestation is contained in the discussion of ecosystem extent, especially in the case studies of Paraguay and Cambodia.
 4.3.2 Invasive species
4.3.2 Invasive species In many cases, it is possible to map the spread of, and impact from, specific invasive species (Van der Meer et al. 2002). There have been considerable advances in using remote sensing to map species that dominate forest canopies, including Surinam cherry or
Melaleuca quinquenervia (McCormick 2002), tamarisk or
Tamarix chinensis (Everitt and Deloach 1990), leucaena or
Leucaena leucocephala (Tsai et al. 2005), maritime pine or Pinus pinaster (Ferreira et al. 2005), Chinese tallow or Sapium sebiferum (Ramsey et al. 2002), and trumpet tree (Cecropia peltata) (Lee et al. 1990).
The majority of plant invasives in native forests occur in the understory where they are often obscured by the canopy. However, where herbaceous weedy plants occur in forest openings and along forest edges and waterways, they are detectable. See, for example, leafy spurge or
Euphorbia esula (Everitt et al. 1995), spotted knapweed or
Centaurea maculosa (Lass et al. 2002), and cogongrass or
Imperata cylindrica (Huang et al. 2001).
Silvicultural practices often enhance the effectiveness of invasives (Richardson 1998; Thysell and Carey 2001). Roads have been shown to be excellent conduits for alien species (Trombulak and Frissell 2000). Adjacent land uses, size of forest fragments, and amount of edge around forest fragments all affect invasion (With 2002). All of these can be monitored to varying degrees using remote sensing. Another valuable use of remote sensing in monitoring invasive species is the effect that some invasives have on forest condition. Working in a Hawaiian montane rain forest, Asner and Vitousek (2005) used an airborne visible and infrared imaging spectrometer from an ER-2 high-altitude aircraft to measure water content and leaf nitrogen (N) concentrations in intact
Metrosideros polymorpha forests and those invaded by
Myrica faya, a nitrogen-fixing exotic tree, and
Hedychium gardnerianum, an understory herb that reduces nitrogen concentration in forest canopies. Bonneau et al. (1999) used remote sensing to classify and track hemlock forests infested by the hemlock woolly adelgid, an exotic insect pest. Monitoring of invasive species is not exclusively a high-resolution endeavor. Bryceson (1991) tracked the Australian plague locust (
Chortoicetes terminiflora) using Landsat TM imagery, and Kharuk et al. (2001) analysed large scale outbreaks of the Siberian moth (
Dendrolimus sibiricus), using AVHRR imagery. (For more on invasive species,
see Chapter 11.)
4.3.3 Climate change – carbon storage and fire Of all the identified drivers of forest biodiversity loss, climate change has the potential to become the leading agent in the coming decades, although its effects remain difficult to predict. Since 1900, the burning of fossil fuels has contributed the most to atmospheric greenhouse gases, although clearing and burning of tropical forests accounts for between 20 and 25 percent of anthropogenic greenhouse gases released each year (Moutinho and Schwartzman 2005). Forests act in concert with climate: influencing it while at the same time being influenced by it. When large areas are left undisturbed and forest growth exceeds harvest, forests can reduce carbon dioxide (CO2) levels in the atmosphere and therefore are important to monitor at the global level. Unfortunately, the capacity of the world’s forests to maintain native composition, structure, and function is being increasingly compromised by the multitude of the numerous stressors they face. With regard to climate-change modelling, remote sensing plays an important and effective role. It provides data on land cover, carbon stocks, rates of change, above-ground biomass, and even some human sources of methane (Rosenqvist et al. 2003).
Predictions concerning the types and severity of changes expected in forest ecosystem composition, structure, and function vary by forest biome and regional history (Watson et al. 1997). Most of the world’s forests are expected to experience some level of disturbance caused by the changing climate, posing a serious threat to global forest biodiversity, forest-based economies, and ecosystem services. A growing body of scientific literature indicates that climate change is already affecting forests by shifting species ranges, fostering pests and pathogens, altering fire disturbance regimes, changing migration patterns, and causing species extinctions. Northern latitude and mountain forest systems are showing the greatest changes. Using time series AVHRR data (1981–1999), trends show a general increase in growing-season length, annual primary productivity, and northward extension of tree line in the Canadian boreal (Mynemi et al. 2001; Zhou et al. 2001). The Canadian boreal has also seen an increase in fire frequency and intensity over recent years (Gillett et al. 2004). In mountain forests, where native forests are also immediately vulnerable to climate change, species changes (including local extinctions) are being reported (Pounds et al. 1999). Monitoring forest ecosystems like tropical cloud forests will require integrating some of the newer remote sensing technologies (e.g., radar) with more-traditional sensors (e.g., Landsat or SPOT), but it could be done in a cost-effective manner because the geographic areas involved are relatively small.
Remote sensing has been shown to be effective at tracking carbon sequestration by forests in a timely and spatially explicit fashion over large regions (Dong et al. 2003), as well as small regions (Turner et al. 2004b), and mapping at both scales plays a substantial role in providing the necessary data for local-, regional-, and global-scale carbon cycle models (Turner et al. 2004a). Remote sensing provides an excellent opportunity to track this important forest function when used in connection with robust carbon mass balance models (Veroustraete and Verstraeten 2004). MODIS has been used frequently to monitor net primary productivity at global and regional scales (Liu et al. 1999), and historical Landsat imagery and JERS-1 SAR are the most useful sensors for establishing a 1990 carbon-stock baseline (Rosenqvist et al. 2003).
A variety of sensors have been used to map and monitor Central African land cover, including carbon sources and sinks (Laporte 2000). The biomass maps that are routinely updated (figure 4.9) provide insight into the changing landscape, which has direct impact on many forms of regional biodiversity. For example, this region is where Africa’s great apes, including mountain gorillas (
Gorilla gorilla beringei), lowland gorillas (
Gorilla gorilla gorilla), and bonobos (
Pan paniscus) occur and where human-induced changes are resulting in rapid declines in many of the species. Monitoring biomass changes (carbon sources and sinks) not only supplies important monitoring data for ecological process models, which have the potential to predict biodiversity losses into the future, but it also provides important monitoring data that can be applied directly today to specific biodiversity concerns.
Forest fires are part of a feedback loop that relates to global climate change. When forests burn, they release carbon dioxide, thus adding to the greenhouse gases and raising the risk of future wildfires. In some parts of the world, where fire suppression has been effective, there has been forest-based fuel buildup. If more forests burn, whether from wildfires or increased prescribed burning, more carbon dioxide will be released back into the atmosphere, where it will join the increased emissions from human causes. Forest fires naturally release tons of carbon dioxide into the atmosphere each year. This is expected to increase in some regions, exacerbating climate change and with direct impacts on local and regional biodiversity. Therefore, mapping and monitoring forest fires are important activities for understanding the role that fires have in impacting climate change and biodiversity. As discussed earlier in this chapter, there is an extensive and ongoing monitoring effort of fires globally, using remote sensing.
4.3.4 Forest degradation A degraded forest is a secondary forest that has lost, through human activities, the structure, function, species composition or productivity normally associated with a natural forest type expected on that site. Hence, a degraded forest delivers a reduced supply of goods and services from the given site and maintains only limited biological diversity. Biological diversity of degraded forests includes many non-tree components, which may dominate in the undercanopy vegetation.
— Ad hoc technical expert group on forest biological diversity,
Convention on Biological Diversity.
Perhaps more than any other region in the world, the Amazon has been the focus of the most studies employing remote sensing to assess forest degradation, which in this region is dominated by selective logging and burning (Nepstad et al. 1999). Using a combination of 1-metre resolution IKONOS data and SPOT 4, Souza et al. (2003) developed a methodology that allowed them to map four classes of forest, including a degraded forest class, defined as either heavily burned or heavily logged and burned, using coarse imagery with accuracy of approximately 86 percent. Grainger (1999) focused on mapping the differences of biomass to map forest degradation.
Regardless of the main focus of mapping forest degradation, it is often important not only to adjust the monitoring frequency to fit the forest type—some forests require multiple surveys per year (e.g., dry tropical forests), while others require less frequent surveys (e.g., temperate coniferous forests)—but also to compile a long time series to differentiate natural variability from human degradation (Lambin 1999). Monitoring forest degradation in a standard way is an ongoing challenge. Remote sensing has been shown to be effective in tracking many characteristics of forests. What is missing are global and national standards for defining what forest degradation activities are most important to track in different regions and then developing a standard methodology so systematic monitoring can be carried out.
The implementation of remote sensing in forest monitoring will require a carefully designed strategy that takes advantage of a wide range of sensors, ancillary data, ground control, and local expertise. Once the data are in hand, it will also be important to find new ways to understand the many cumulative and synergistic threats to the world’s forests so that meaningful solutions can be designed and implemented.

4.4 Data and Other Resources
Global Forest Resources Assessment (FRA 2000 and FRA 2005) The UN Food and Agricultural Organization, at the request of its member nations and the world community, has regularly reported on the state, changes, and conditions of the world’s forests. The
Global Forest Resources Assessment (FRA 2005) provides crucial information describing the state and conditions of forest resources for 2005 and as well as changes over the past 15 years. It is available on FAO’s website along with past assessments.
The global forest map is one of the many outputs produced by these reports. The forest map in the 2000 report was produced from the Global Land Cover Characteristics Database (GLCCD), a land cover data set at 1-kilometre spatial resolution derived from AVHRR satellite images. In addition to a map showing the distribution of forests, maps of forests by ecological zones were also prepared.
The forest map for the next assessment (
FRA 2010) will be based on MODIS 250 m resolution data.
For forest maps see:
www.fao.org/forestry/site/24815/en http://edcsns17.cr.usgs.gov/glcc/fao/index.html FRA 2005
Global Observation of Forest and Land Cover Dynamics The stated objective of the
Global Observation of Forest and Land Cover Dynamics (GOFC-GOLD) is to improve the quality and availability of observations of forests at regional and global scales and to produce useful, timely, and validated information products.
Tropical Rain Forest Information Center The
Tropical Rain Forest Information Center was established to provide data, products, and information services to NASA. Landsat and other high resolution satellite remote sensing data, as well as digital deforestation maps and databases, are available online.
Global Forest Watch Global Forest Watch is an inititative of the World Resources Institute. It provides numerous forest maps (including atlases) and assessments online for various regions of the world, produced in collaboration with GFW partners in Brazil, Canada, Central Africa, Chile, Indonesia, Russia, Untited States and Venezuela.
Global Fire Data The University of Maryland provides historic and nearly real-time fire ignition data based on MODIS imagery at the global, regional, and national levels. Early-warning system details can be obtained from the
Global Fire Monitoring Center (GFMC).
Global Forest Fragmentation Data Digital forest fragmentation maps derived from Global Land Cover Characterization (GLCC) land cover maps (based on Advanced Very High Resolution Radiometer AVHRR, circa 1992) are available online. Each pixel value represents a forest fragmentation category for the surrounding 81square kilometre landscape. The maps are directly comparable to the GLCC maps and are distributed by continent in the GLCC format.
National and Regional Forest Data Resources Brazilian Amazon Tropical Rain Forest Information Center provides numerous maps in pdf format for the region, including deforestation, forest cover, and forest classification.
Amazon Forest Inventory Network (RAINFOR) is an international network established to monitor the biomass and dynamics of Amazonian forests.
Canada Natural Resources Canada (NRCAN) provides numerous national forest themes and images.
Central Africa
Global Land Cover Facility provides forest change data for central Africa.
Global Forest Watch provides forest assessment and atlas data for central Africa, particularly in Cameroon.
Central America Land Cover
Center for International Earth Science Information Network (CIESIN) is a centre within the Earth Institute at Columbia University in the United States. CIESIN provides recent Central American land cover data online.
SERVIR is a regional visualization and monitoring system for Mesoamerica that integrates satellite and other geospatial data and makes them widely available.
Europe
European Forest Institute is an independent nongovernmental organization conducting European forest research and providing numerous forest data layers for Europe.
European Forest Information Scenario Model (EFISCEN) collects and provides national forest inventory data for 30 European nations, plus parts of Russia.
India Forest Survey of India assesses forest cover of the country on a two-year cycle, using satellite data. The main objectives are to present the information on Indian forest resources at state and district levels and to prepare forest cover maps on a 1:50,000 scale.
Southeast Asia Tropical Rain Forest Information Center (TFRIC) provides numerous maps in pdf format for the region, including deforestation, forest cover, and forest classification.
4.5 References
Achard, F., H. D. Eva, P. Mayaux, H.-J. Stibig, and A. Belward. 2004. Improved estimates of net carbon emissions from land cover change in the tropics for the 1990s. Global Biogeochemical Cycles 18: ART 91804.
Achmaliadi, R., I.G. Maha Adi, Y.M. Hardiono, H. Kartodihardjo, F.C.H. Malley, D.A. Mampioper, E.G.T. Manurung, A. Nababan, L.B. Pangkali, A. Ruwindrijarto, L.L.M. Situmorang, and Wardiyono. 2002. The state of the forest: Indonesia. Global Forest Watch. Washington, D.C.
http://www.globalforestwatch.org/common/indonesia/sof.indonesia.english.low.pdf. (accessed January, 2007).
Aguilar, A. 2005. Remote sensing of forest regeneration in highland tropical forests. GIScience and Remote Sensing 42(1): 66-79.
Aksenov, D., M. Karpachevskiy, S. Lloyd, and A. Yaroshenko. 1999. The last of the last: The old-growth forests of boreal Europe. Taiga Rescue Network.
www.globalforestwatch.org/english/about/publications.htm. (accessed January, 2007).
Aksenov, D., et al. 2002. Atlas of Russia’s intact forest landscapes. Global Forest Watch, World Resources Institute. Moscow, Russia.
www.globalforestwatch.org/english/about/publications.htm. (accessed January, 2007).
Aksenov, D., M. Yu. Dubinin, M.L. Karpachevskiy, N.S. Liksakova, V.E. Skvortsov, D. Yu. Smirnov, and T.O. Yanitskaya. 2006. Mapping high conservation value forests of Primorsky Kray, Russian Far East. Global Forest Watch Russia.
www.globalforestwatch.org/english/about/publications.htm. (accessed January, 2007).
Anger, C.D., S. Mah, and S.K. Babey. 1994. Technological enhancements to the compact airborne spectrographic imager (casi). Proceedings of the First International Airborne Remote Sensing Conference and Exhibition. Ann Arbor, Michigan: Environmental Research Institute of Michigan. 2: 205-213.
Asner, G.P., and P.M. Vitousek. 2005. Remote analysis of biological invasion and biogeochemical change. Ecology 102(12): 4383-4386.
Barreto, P., C. Souza Jr., R. Noguerón, A. Anderson, and R. Salomão. 2006. Human pressure on the Brazilian amazon forests. Amazon Institute of People and the Environment and World Resources Institute 2006. Washington, D.C.
www.globalforestwatch.org/english/about/publications.htm. (accessed January, 2007).
Bartholomé, E., and A.S. Belward. 2005. GLC2000: A new approach to global land cover mapping from Earth observation data. International Journal of Remote Sensing 26(9): 1959-1977.
Bevilacqua, M., Cárdenas, L., Flores, A.L., Hernández, L., Lares B., E., Mansutti R., A., Miranda, M., Ochoa G., J., Rodríguez, M. and E. Selig. 2002.The State of Venezuela’s Forests. A Case Study of the Guayana Region. A Global Forest Watch Report, World Resources Institute, Washington D.C., 132 pages.
Bonneau, L.R., K.S. Shields, and D.L. Civco. 1999. Using satellite images to classify and analyse the health of hemlock forests infested by the hemlock woolly adelgid. Biological Invasions 1:55-267.
Bryant, D., D. Nielsen, and L. Tangley. 1997. The last frontier forests: Ecosystems and economies on the edge. World Resources Institute. Washington, D.C.
Bryceson, K.P. 1991. Likely locust infestation areas in western New South Wales, Australia, located by satellite. Geocarto International 6(4): 21-37.
Clark, D.B., J.M. Read, M.L. Clark, A.M. Cruz, M.F. Dotti, and D.A. Clark. 2003. Application of 1-m and 4-m resolution satellite data to ecological studies of tropical rain forests. Ecological Applications 14(1): 61-74.
Cohen, W.B., Spies, T.A., and M. Fiorella. 1995. Estimating the age and structure of forests in a multi-ownership landscape of western Oregon U.S.A. International Journal of Remote Sensing 16(4): 721-746.
Cohen, W.B., T.K. Maiersperger, T.A. Spies, and D.R. Oetter. 2001. Modelling forest cover attributes as continuous variables in a regional context with Thematic Mapper data. International Journal of Remote Sensing 22: 2279-2310.
Coops, N.C., R.H. Waring, and J.J. Landsberg. 1998. Assessing forest productivity in Australia and New Zealand using a physiologically-based model driven with averaged monthly weather data and satellite-derived estimates of canopy photosynthetic capacity. Forest Ecology and Management 104: 113-127.
DeFries, R., F. Achard, S. Brown, M. Herold, D. Murdiyarso, B. Schlamadinger, and C. de Souza Jr. 2006. Reducing greenhouse gas emissions from deforestation in developing countires: Considerations for monitoring and measuring. GOFC-GOLD Rome, Italy.
Donahey, J. GIS and remote sensing analyses of forest plantations in Costa Rica’s Atlantic Lowland Area.
www.nrem.iastate.edu/ECOS/weblogs/Donahey_FinalReport2006.pdf. (accessed January, 2007).
Dong, J., R.K. Kaufmann, R.B. Myneni, C.J. Tucker, P.E. Kauppi, J. Liski, W. Buermann, V. Alexeyev, and M.K. Hughes 2003. Remote sensing estimates of boreal and temperate forest woody biomass: Carbon pools, sources, and sinks, Remote Sensing of Environment 84: 393-410.
Drake, J.B., R.O. Dubayah, D.B. Clark, R.G. Knox, J.B. Blair, M.A. Hofton, R.L. Chazdon, J.F. Weishampel, and S.D. Prince. 2002. Estimation of tropical forest structural characteristics using large-footprint lidar. Remote Sensing of Environment 79: 305-319.
Evans, J., N. Pelkey, and D. Haskell. 2002. An assessment of forest change on the Cumberland Plateau in Southern Tennessee. Report to the US EPA and USF&WS. lal.sewanee.edu/research/assessment.html. (accessed January, 2007).
Everitt, J.H., and C.J. Deloach. 1990. Remote sensing of Chinese tamarisk (Tamarix chinensis) and associated vegetation. Weed Science 38(3): 273-278.
Everitt, J.H., G.L. Anderson, D.E. Escobar, M.R. Davis, N.R. Spencer, and R.J. Andrascik. 1995. Use of remote sensing for detecting and mapping leafy spurge (
Euphorbia esula). Weed Technology 9(3): 599-609.
FAO (Food and Agriculture Organization). 2006. Expert Consultation on global forest resources assessment: towards FRA 2010: Kotka V. Kotka, Finland.
www.metla.fi/tapahtumat/2006/kotkav/index.htm.
Ferreira, M.T., F.C. Aguiar, and C. Nogueira. 2005. Changes in riparian woods over space and time: Influence of environment and land use. Forest Ecology and Management 212: 145-159.
FRA 2000 . Global Forest Resources Assessment 2000: Main Report. Food and Agrilculture Organization. Rome, Italy.
www.fao.org/forestry/site/fra2000report/en/.
FRA 2005. Global Forest Resources Assessment 2005: Progress towards sustainable forest management. Food and Agriculture Organization. Rome, Italy.
www.fao.org/docrep/008/a0400e/a0400e00.htm. (accessed January, 2007).
Franklin, S.E., M.A. Wulder, and G.R. Gerylo. 2001. Texture analysis of IKONOS panchromatic imagery for Douglas-fir forest age class separability in British Columbia. International Journal of Remote Sensing 22: 2627-2632.
Fraser, R.H., Z. Li, and J. Cihlar. 2000. Hotspot and NDVI differnecing synergy (HANDS): A new technique for burned area mapping over boreal forest. Remote Sensing of Environment 74: 362-376.
Friedl, M.A., D.K. McIver, J.C.F. Hodges, X.Y. Zhang, D. Muchoney, A.H. Strahler, C.E. Woodcock, S. Gopal, A. Schneider, A. Cooper, A. Baccini, F. Gao, and C. Schaaf. 2002. Global land cover mapping from MODIS: Algorithms and early results. Remote Sensing of Environment 83(1-2): 287-302.
Friedl, M.A. 2007. Validation of the consistent-year v003 MODIS land cover product.
www-modis.bu.edu/landcover/userguidelc/consistent.htm. (accessed January, 2007).
Gillett, N.P., Weaver, A.J., Zwiers, F.W. and M.D. Flannagan. 2004. Detecting the effect of human-induced climate change on Canadian forest fires. Geophysical Research Letters 31 (18): L18211.
GOFC-GOLD. 2004. CEOS land cover validation workshop. Boston, MA. 2-5 February 2004.
Grainger, A. 1999. Constraints on modelling the deforestation and degradation of tropical open woodlands. Global Ecology and Biogeography 8(3-4): 179-190.
Greenpeace. 2006. Roadmap to recovery: The world’s last Intact forest landscapes.
www.intactforests.org/publications/publications.htm. (accessed January, 2007).
Grégoire, J., K. Tansey, and J. Silva. 2003. The GBA2000 initiative: Developing a global burnt area database from SPOT-VEGETATION imagery. International Journal of Remote Sensing 24(6): 1369-1376.
Hansen, M., R.S. DeFries, J.R.G. Townshend, M. Carroll, C. Dimiceli, and R.A. Sohlberg. 2003. Global percent tree cover at a spatial resolution of 500 meters: First results of the MODIS vegetation continuous fields algorithm.”Earth Interactions 7(10): 1-15.
Hansen, M., and R. DeFries. 2004. Detecting long term forest change using continuous fields of tree cover maps from 8km AVHRR data for the Years 1982–99. Ecosystems 7: 695–716.
Heilman, Jr. G.E., J.R. Strittholt, N.C. Slosser, and D.A. DellaSala. 2002. Forest fragmentation of the conterminous United States: Assessing forest intactness through road density and spatial characteristics. Bioscience 52(5): 411-422.
Helmer, E.H., S. Brown, and W.B. Cohen. 2000. Mapping montane tropical forest successional stage and land use with multi-date Landsat imagery. International Journal of Remote Sensing 21(11): 2163-2183.
Holling, C.S. 1992. Cross-scale morphology, geometry, and dynamics of ecosystems. Ecological Monographs 62:447-502.
Huang, Y., M. Bruce, L. Byrd, and J. Mask. 2001. Using wavelet transform of hyperspectral reflectance curves for automated monitoring of Imperata cylindrical (cogongrass). GEOScience and Remote Sensing 5: 2244-2246.
Hunter, M.L. 1999. Maintaining Biodiversity in Forest Ecosystems. Cambridge University Press. New York NY., 1999.
Jensen, J.R., F. Qiu, and M. Ji. 1999. Predictive modelling of coniferous forest age using statistical artificial neural network approaches applied to remote sensor data. International Journal of Remote Sensing 20(14): 2805-2822.
Jiang, H., J.R. Strittholt, P.A. Frost, and N.C. Slosser. 2004. The classification of late seral forests in the Pacific Northwest, USA using Landsat ETM+ imagery. Remote Sensing of Environment 91: 320-331.
Justice, C.O., L. Giglio, S. Korontzi, J. Owens, J.T. Morisette, D. Roy, J. Descloitres, S. Alleaume, F. Petitcolin, and Y. Kaufman. 2002. The MODIS fire products. Remote Sensing and Environment 83: 244-262.
Kerr, J.T., and M. Ostrovsky. 2003. From space to species: Ecological applications for remote sensing. Trends in Ecology and Evolution 18(6): 299-314.
Kharuk, V.I., A.A. Kozhukhovskaya, I.A. Pestunov, K. Ranson, and G.M. Tsibul’skiy. 2001. Use of NOAA/AVHRR imagery in monitoring Siberian moth outbreaks. Mapping Sciences and Remote Sensing 38(4): 272-281.
Kimes, D.S., R.F. Nelson, D.L. Skole, and W.A. Salas. 1998. Accuracies in mapping secondary tropical forest age from sequential satellite imagery. Remote Sensing and Environment 65(1): 112-120.
Lambin, E.F. 1999. Monitoring forest degradation in tropical regions by remote sensing: some methodological issues. Global Ecology and Biogeography 8(3-4): 191-198.
Lammertink, J.M., J.A. Rojas-Tome, F.M. Casillas-Orona, and R.L. Otto. 2007. Estatus y conservación de los bosques maduris y la saves endémicas en la zona de pino-encino de la Sierra Madre Occidental, México.
www.worldwildlife.org/bsp/publications/lac/status/status1.html#methods. (accessed January, 2007).
Laporte, N. 2000. An integrated forest monitoring system for Central Africa.
lcluc.umd.edu/products/pdfs/GOFC/GOFC-LaPorte2000.pdf. (accessed January, 2007).
Lass, L.W., D.C. Thill, B. Shafii, and T.S. Prather. 2002. Detecting spotted knapweed (
Centaurea maculosa) with hyperspectral remote sensing technology. Weed Technology 16(2): 426-432.
Lee, D.W., R.A. Bone, S.L. Tarsis, and D. Storch. 1990. Correlates of leaf optical properties in tropical forest sun and extreme-shade plants. American Journal of Botany 77(3): 370-380.
Lee, P., D. Aksenov, L. Laestadius, R. Noguerón, and W. Smith. 2003. Canada’s large intact forest landscapes. Global Forest Watch Canada and World Resources Institute. Washington, DC.
www.globalforestwatch.org/english/about/publications.htm. (accessed January, 2007).
Lee, P., J.D. Gysbers, and Z. Stanojevic. 2006. Canada’s forest landscape fragments: A first approximation. Global Forest Watch Canada. Edmonton, Alberta. 2006.
www.globalforestwatch.ca/publications_and_maps.htm. (accessed January, 2007).
Lefsky, M.A., W.B. Cohen, G.G. Parker, and D.J. Harding. 2002. Lidar remote sensing for ecosystem studies. Bioscience 52(1): 19-30.
Liu, J., J.M. Chen, J. Cihlar, and W. Chen. 1999. “Net primary productivity distribution in the BOREAS region from a process model using satellite and surface data.” Journal of Geophysical Research 104.D22: 27735-27754.
Loveland, T.R., B.C. Reed, J.F. Brown, D.O. Ohlen, J. Zhu, L.Yang, and J.W. Merchant. 2000. Development of a global land cover characteristics database and IGBP DISCover from 1-km AVHRR data. International Journal of Remote Sensing 21(6/7): 1303-1330.
Mayaux, P., and E.F. Lambin. 1997. Tropical forest area measured from global land-cover classification: Inverse calibration models based on spatial textures. Remote Sensing of Environment 59: 29-43.
Mayaux, P., A. Strahler, H. Eva, M. Herold, A. Shefali, S. Naumov, S. Dorado, C. Di Bella, D. Johansson, C. Ordoyne, I. Kopin, L. Boschetti, and A. Belward. 2006. Validation of the global land cover 2000 map. IEEE-Transactions on Geoscience and Remote Sensing 44(7): 1728-1739.
McCormick, C.M. 2002. Mapping exotic vegetation in the Everglades from large-scale aerial photographs. Photogrammetric Engineering and Remote Sensing 65(2): 179-184.
McIver, D.K., and M.A. Friedl. 2002. Using prior probabilities in decision-tree classification of remotely sensed data. Remote Sensing of Environment 81(2-3): 253-261.
Minnemeyer, S. 2002. An analysis of access into central Africa’s rainforests. Global Forest Watch. Washington, DC.
www.globalforestwatch.org/english/about/publications.htm. (accessed January, 2007).
Mollicone, D., F. H. Achard, A. Eva, S. Belward, A. Federici, V.C. Lumicisi, H. Risso, Stibig, and R. Valentini. 2003. Land use change monitoring in the framework of the UNFCCC and its Kyoto Protocl: Report on current capabilities of satellite remote sensing technology. European Communities, Luxembourg, 2003.
Moran, M.S., T.R. Clarke, Y. Inoue, and A. Vidal. 1994. Estimating crop water deficit using the relation between surface-air temperature and spectral vegetation index. Remote Sensing and Environment 49: 246-263.
Moutinho, P., and S. Schwartzman. 2005. Tropical deforestation and climate change. Amazon Institute of Environmental Research.
Myneni R.B., Dong J., Tucker C.J., Kaufmann R.K., Kauppi P.E., Liski J., Zhou L., Alexeyev V., and M.K. Hughes. 2001. A large carbon sink in the woody biomass of Northern forests. Proceedings of the National Academy of Sciences of the USA 98: 14784–14789.
Neira, E., H. Verscheure, and C. Revenga. 2002. Chile’s frontier forests: Conserving a global treasure. Global Forest Watch,. Washington,D.C.
www.globalforestwatch.org/english/about/publications.htm. (accessed January, 2007).
Nelson, T., K.O. Niemann, and M. Wulder. 2003. Spatial statistical techniques for aggregating point objects extracted from high spatial resolution remotely sensed imagery. Journal of Geographical Systems 4: 423-433.
Nepstad, D.C., J.A. Verissimo, A. Alencar, C. Nobre, E. Lima, P. Lefebvre, P. Schlesinger, C. Potter, P. Moutinho, E. Mendoza, M. Cochrane, and V. Brooks. 1999. Large-scale impoverishment of Amazonian forests by logging and fires. Nature 398: 505-508.
Noguerón, R. 2002. Low-access forests and their level of protection in North America. World Resources Institute. Washington, D.C.
www.globalforestwatch.org/english/about/publications.htm. (accessed January, 2007).
Ohmann, J.L. and Gregory, M.J. 2002. Predictive mapping of forest composition and structure with direct gradient analysis and nearest-neighbor imputation in coastal Oregon, U.S.A. Canadian Journal of Forest Research 32: 725-741.
O’Neill, R.V., et al. 1986. A hierarchical concept of ecosystems. Princeton University Press.
Ramsey, E.W., G.A. Nelson, S.K. Sapkota, E.B. Seeger, and K.D. Martella. 2002. Mapping Chinese tallow with color-infrared photography. Photogrammetric Engineering and Remote Sensing 68(3): 251-255.
Ratnayake, R.L. 2006. Identification of age groups of managed pine plantation using remote sensing data.
www.gisdevelopment.net/proceedings/mapindia/2006/envi.htm. (accessed January, 2007).
Richardson, D.M. 1998. Forestry trees as invasive species. Conservation Biology 12(1): 18-26.
Riitters, K., J. Wickham, R. O’Neill, B. Jones, E. Smith, J. Coulston, T. Wade, and J. Smith. 2002. Fragmentation of continental United States forests. Ecosystems 5(8): 815-822.
Riitters, K., J. Wickham, R. O’Neill, B. Jones, and E. Smith. 2004. Global-scale patterns of forest fragmentation. Conservation Ecology 4(2): 3.
http://www.consecol.org/vol4/iss2/art3/.
Rosenqvist, A., A. Milne, R. Lucas, M. Imhoff, and C. Dobson. 2003. Environmental Science and Policy 6: 441-455.
Smith, W., B. Evans, T. Wilson, A. Nikiforuk, K. Baltggailis, A. Brady, L. Brewster, T Gray, W. Horter, P. Lee, R. Livernash, C. McGlynn, S. Minnemeyer, A. O’Carroll, A. Penn, G. Quaile, M. Sawyer, E. Selig, and D. Bryant. 2000. Canada’s forests at a crossroads: An assessment in the year 2000. World Resources Institute, Washington, D.C.
www.globalforestwatch.org/english/about/publications.htm. (accessed January, 2007).
Souza, Jr. C., L. Firestone, L.M. Silva, and D. Roberts. 2003. Mapping forest degradation in the Eastern Amazon from SPOT4 through spectral mixture models. Remote Sensing of Environment 87: 494-506.
Strahler, A.H., L. Boschetti, G.M. Foody, M. Friedl, M. Hansen, M. Herold, P. Mayaux, J. Morisette, S. Stehman, and C.E. Woodcock. 2006. Global land cover validation: Recommendations for evaluation and accuracy assessment of global land cover maps. European Communities, Luxembourg.
Steininger, M.K. 1996. Tropical secondary forest regrowth in the Amazon: Age, area and change estimation with Thematic Mapper data. International Journal of Remote Sensing 17(1): 9-27.
Steininger, M.K., C.J. Tucker, J.R.G. Townshend, T.J. Killeen, A. Desch, V. Bell, and P. Ersts. 2001. Tropical deforestation in the Bolivian Amazon. Environmental Conservation 28: 127-134.
Steyaert, L.T., F.G. Hall, and T.R. Loveland. 1997. Land cover mapping, fire regeneration, and scaling studies in the Canadian boreal forest with 1km AVHRR and Landsat TM data. Journal of Geophysical Research 102.D24: 581-598.
Strittholt, J.R., R. Noguerón, M. Alvarez, and J. Bergquist. 2006. Mapping undisturbed landscapes in Alaska: An overview report. Conservation Biology Institute and World Resources Institute.
www.globalforestwatch.org/english/about/publications.htm (accessed January, 2007).
Tansey, K., J-M. Grégoire, D. Stroppiana, A. Sousa, J.M.N. Silva, J.M.C. Pereira, L. Boschetti, M. Maggi, P.A. Brivio, R. Fraser, S. Flasse, D. Ershov, E. Binaghi, and D. Graetz. 2004. Vegetation burning in the year 2000: Global burned area estimates from SPOT VEGETATION data. Journal of Geophysical Research 109.D14: CiteID D14S03.
Thysell, D.R. and A.B. Carey. 2001. Manipulation of density of Psudotsuga menziesii canopies: preliminary effects on understory vegetation. Canadian Journal of Forest Research 31(9): 1513-1525.
Treuhaft, R.N., B.E. Law, and G.P. Asner. 2004. Forest attributes from radar interferometric structure and its fusion with optical remote sensing. Bioscience 54: 561-571.
Trichon, V., D. Ducrot, and J.P. Gastellu-Etchegorry. 1999. SPOT4 potential for the monitoring of tropical vegetation: A case study in Sumatra. International Journal of Remote Sensing 20(14): 2761-2785.
Trombulak, S.C. and C.A. Frissell. 2000. Review of ecological effects of roads on terrestrial and aquatic communities. Conservation Biology 14(1): 18-30.
Trotter, C.M., J.R. Dymond, and C.J. Goulding. 1997. Estimation of timber volume in a coniferous plantation forest using Landsat TM. International Journal of Remote Sensing 18(10): 2209-2223.
Tsai, F., E. Lin, and H. Wang. 2005. Detecting invasive plant species using hyperspectral satellite imagery. GEOScience and Remote Sensing Symposium, 2005, IGARSS’05 Proceedings. 2005 IEEE International 4:3002-3005.
Tucker, C. J., and J.R.G. Townshend. 2000. Strategies for tropical forest deforestation assessment using satellite data. International Journal of Remote Sensing, 21: 1461-1472.
Turner, D.P., S.V. Ollinger, and J.S. Kimball. 2004a. “Integrating remote sensing and ecosystem process models for landscape to regional scale analysis of the carbon cycle.” Bioscience 54(6): 573-584.
Turner, D.P., M. Guzy, M.A. Lefsky, W.D. Ritts, S. van Tuyl, and B.E. Law. 2004b. “Monitoring forest carbon sequestration with remote sensing and carbon cycle modelling.” Environmental Management 33(4): 457-466.
UNEP World Conservation Monitoring Centre. 2007. Global Distribution of Current Forests.
http://www.unep-wcmc.org/forest/global_map.htm (accessed June 2007)
Van der Meer, F., Schmidt, K.S., Bakker, A., & Bijker, W., 2002. New Environmental RS systems. In: Environmental Modelling with GIS and RS. Skidmore, A. K., editor, Taylor & Francis, London, pp. 26-51.
Van de Pol, M.J., L.A. Nsoyuni, S. Minnemeyer, and R. Ngoufo. 2005. Interactive forestry atlas of Cameroon (version 1.0): An overview. Global Forest Watch. Washington, DC.
www.globalforestwatch.org/english/about/publications.htm. (accessed January, 2007).
Veroustraete, F., and W.W. Verstraeten. 2004. European forest carbon mass balance estimated with remote sensing and the production efficiency model C-fix: A hot future unfolds for Kyoto Protocol implementation. American Geophysical Union, Fall Meeting 2004.
adsabs.harvard.edu/abs/2004AGUFM.B43C..06V. (accessed January, 2007).
Watson, R.T., M.C. Zinyowera, R.H. Moss, and D. Dokken. 1997. IPCC special report on the regional impacts of climate change: An assessment of vulnerability. Intergovernmental Panel on Climate Change.
www.grida.no/climate/ipcc/regional/198.htm. (accessed January, 2007).
With, K.A., Pavuk, D.M., Worchuck, J.L., Oates, R. K., Fisher, J.L. 2002. Threshold Effects of Landscape Structure on Biological Control in Agroecosystems. Ecological Applications 12 (1): 52-65.
Wulder, M.A., R.J. Hall, N.C. Coops, and S.E. Franklin. 2004. High spatial resolution remotely sensed data for ecosystem characterization. Bioscience 54(6): 511-521.
Yaroshenko, A. 2001. The last intact forest landscapes of Northern European Russia. Greenpeace Russia and Global Forest Watch.
Zhou, L., C.J. Tucker, R.K. Kaufmann, D. Slayback, N.V. Shabanov, I. Fung, and R.B. Myneni. 2001. “Variations in northern vegetation activity inferred from satellite data and vegetation index during 1981-1999.” Journal of Geophysical Research 106: 20069-20083.